

Autopoiesis - life maintaining and reproducing itself
I am actively working on this page. Updated 01-03-21 to include Hofmeyr's contribution and repair damage to page (somehow the bottom part got deleted).Introduction
Here we look at one of the most important theories in biology, since it goes a long way to explaining what life really is and how it works, but unlike other foundational theories, such as Darwin and Wallace's theory of evolution by natural selection, this one has been largely neglected and remains in a sort of intellectual ghetto. I hope to contribute to its wider appreciation, though I suppose I am also in the ghetto now.
The word autopoiesis is a pseudo-Greek combination of 'auto-' meaning 'self' and 'poiesis', meaning 'creation'. This is good because it means the ability of a system to construct its own component parts and self-organise these into a co-ordinated whole and maintain itself without help from another system. You might ask, with incredulity, is it possible for something to create itself? Well, we all do it all the time, indeed this is a defining feature of life. The reason is that every cell in the body is constantly having to remake itself: a lot of the amino acids, sugars and lipids absorbed by a bacterium, or e.g. an epithelial cell in your skin, is used to manufacture the component parts of that same cell. Sure living cells continue by growing and dividing into daughter cells, but this process is itself only an extension of self making (when there is sufficient surplus of component parts to make a copy, then the cell can reorganise into two duplicates and (very delicately) reproduce itself. The critical part of self-making is not the manufacture of materials (e.g. enzymes and scaffold proteins), or even molecular machines (such as protein pumps or the double-motor ATP synthasase complex), it is the correct organisation of these in relation to one another, which has to happen at the same time as they are working. According to the theory of autopoiesis, almost the entire dynamic organisation of the cell is devoted to this activity: a basic cell is the system that is needed for continual self-creation, i.e. no other system will do it.
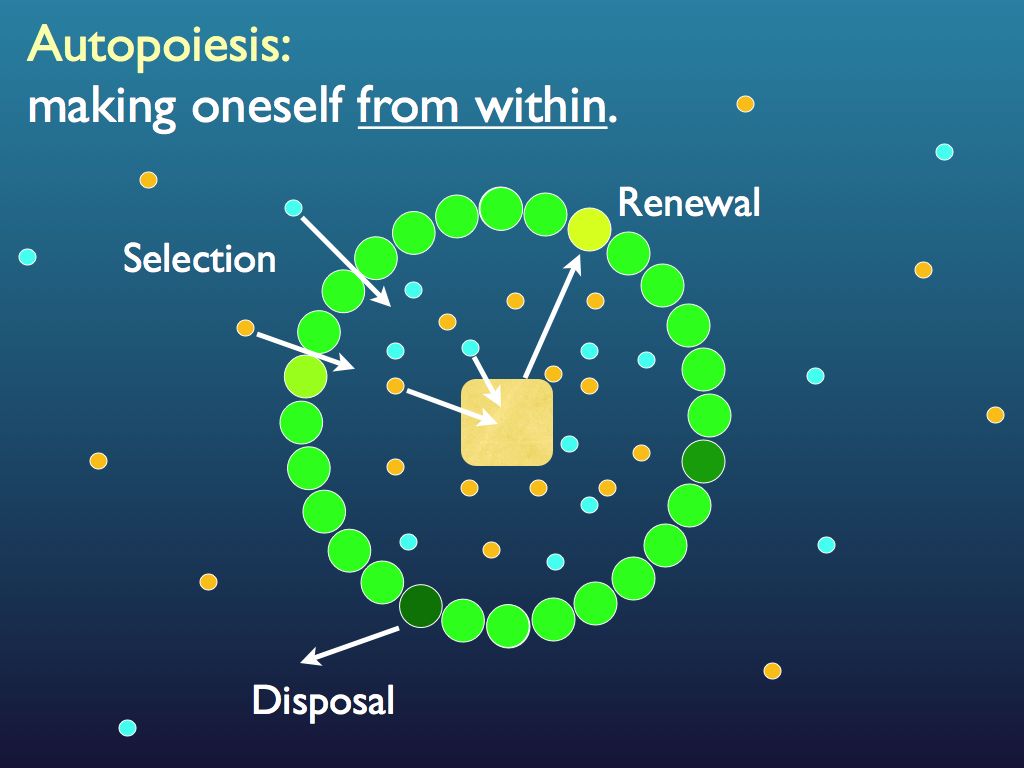
Diagram based on a figure by Pier Luigi Luisi (see below).
We say 'almost' and 'basic cell', because of course there are plenty of cells that produce and process materials, or take other actions that are not necessary for autopoiesis, for example in the processing of lipids in the liver, and the production of antibodies by immune cells, or the conducton of action potentials in nerve cells. A basic cell is the more or less abstract concept of one stripped down to the minimum necessary to persist longer than its component parts. To do that, it has to perform autopoiesis, but nothing else.
The diagram above illustrates the essential features of this. A cell has to select the right materials (molecules) from its environment and contain them in the right concentration within. This sets a requirement for a containing boundary: the tegument that in practice consists of at least a permeable membrane with material filtering subsystems embedded within it (see below). This tegument, as with all complicated molecular structures, breaks down under all the environmental assaults it receives as well as through the inevitable decay dictated by the second law of thermodynamics. For this reason it must be continually repaired, the simplest way to do that being to replace damaged components. The replacement parts (molecular structures) are made form the raw materials selected from the environment by a synthesis system. This system is also manufactured from the raw materials selected from the environment and is itself subject to breakdown and decay. It therefore has to re-manufacture its own component parts, whilst it is still working. Indeed part of its job in working is precisely the remaking of its own components and the correct arrangement of these to form its own structure. That, folks, is autopoiesis. It is good to recognise at this point that we are dealing with three 'elements of physical reality': materials (their composition and concentrations at the molecular level); organisation (which we know to be embodied information) and process (the dynamics of continual renewal that here consists of organised chemical reactions).
Some non-living things show how weird autopoiesis really is
A vortex, such as a whirlpool is self-organising and constructs itself, in a sense, but does not qualify as autopoietic because its component parts are really just particles in fluid phase and the vortex does nothing to make them that way. We might one day produce an 'autonomous' robot that can construct itself by assembling basic components and if it were to exist, then it would arguably be autopoeitic. To achieve the task, though, it would need to somehow exist in some very simple form to begin with and boot-strap itself into full form, otherwise it would not be making itself. This is very hard to conceive, not least because it involves circular causation: a thing being the cause of itself. If we relax the requirements a little, we might imagine that it is only required to make copies of itself from assembling basic components. In this case, it would have to perceive the components available and have the information necessary for assembling them in the right way. So far, the only systems physically capable of this are living systems: every time a cell divides, in a sense a robot creates a copy of itself. We do not know of anything other than living systems that can do this, though we still don't really understand how they first emerged (presumably by boot-strap self creation) (see our Origins Theme). On the other hand, we do now have a good theory for how bootstrap self-creation works in abstract terms of causal relationships. This is explained by Robert Rosen's theory of Relational Biology that I highlight on the causation webpage and it is well worth reading in conjunction with this page: more on that shortly.
Maturana and Varela - radical biology
The theory of autopoiesis (self making and self maintenance) was originally conceived with a single cell in mind, by the Chilean scientists Humberto Maturana and Francisco Varela who published it in two monographs (1980 and 1988). The term itself was first used in 1972 by Maturana, in response, he says, to this thought "If indeed the circular organization is sufficient to characterize living systems as unities, then one should be able to put it in more formal terms." (quoted from Maturana & Varela, 1980, p. xvii). The intention has always been to define life as a common propery of living systems as a whole, as opposed to merely listing features that we observe living systems to hold in common (such as reproduction and metabolism). Accordingly, their definition of life became "autopoiesis in the physical space"; that is the process of autopoieses enacted (or realised) by a physical system.

At the heart of the idea of autopoiesis is the special organisation of a system that works to maintain its own organisation (hence the circularity referred to by Maturana).
An early formal definition was:
"autopoiesis is attributed to a machine (delineated as a a network of processes) which through that network of processes produces the components that:
"(1) through their
interactions and transformations continuously regenerate and realize
the network of processes (relations) that produced them; and
(2) constitute it (the machine) as a concrete unity in the space in which they [the components] exist by specifying the topological domain of its realization as such a network."
(2) constitute it (the machine) as a concrete unity in the space in which they [the components] exist by specifying the topological domain of its realization as such a network."
(Varela, 1979, p. 13), as quoted in Encyclpaedia Autopoietica (Whitaker, 2003).
The term machine is important in this context (and specifically different from that railed against by Rosen).
Quoting Varela:
"In saying that living systems are
'machines' we are pointing to several notions that should be made
explicit. First we imply a nonanimistic view ... Second, we are
emphasizing that a living system is defined by its organization, and
hence that it can be explained as any organization is explained, that
is, in terms of relations, not of component properties. Finally, we are
pointing out from the start the dynamism apparent in living systems and
which the word 'machine' or 'system' connotes."
Later, they reformulated autopoiesis as “a closed topological space that ‘continuously generates and specifies its own organization through its operation as a system of production of its components, and does this in an endless turnover of components.”
This definition further emphasises the organisational property of autopoiesis, so is more compatible with Rosen's (M,R)-system theory (though achieving that was not their purpose).
Subsequently Maturana and Varela have approved the extension of autopoiesis beyond the cell since "the basic requirements for life, of homeostasis and growth, are true also for the community of cells in an organism and for a community of organisms in an ecosystem". It seems a small step in relation to a multi-cellular organism, but is highly contentious to claim this of ecological communities (though some progress has been made in presenting evidence at that level (see theoretical developments by e.g. Gatti et al. 2017 and Farnsworth et al. 2017, with evidence such as the community organisation of sponges (Moitinho-Silva 2017) and microbial community organisation (e.g. Lu et al 2018)). Essentially for its authors, autopoiesis is a definition of life, so that any system which is autopoietic is, by their reasoning, living.
A system may be considered autopoietic if the parts to the organization interact with each other in such a manner that they are continuously producing and maintaining the pattern and elementary parts that constitute the system. Clearly this definition has a close relationship with the autocatalytic sets of Hordijk, Steel and Kauffman and also with the formal definitions (especailly (M,R)-Systems) of Rosen.
Relation to Autonomy
Autopoiesis is part of the set of necessary conditions for Autonomy because it is a way of ensuring causal closure, though in theory that may be achieved in a more general way (see the description of Rosen's (M,R) systems). Living systems are both (simultaneously) separate and integrated with the rest of the world. They are separate only because they are organisationally closed so have their own source of causation (as the (M,R)-system theory explains). It is this organisational closure that gives them their identity, otherwise they could not, in principle, be separated from the rest of the world. But they are integrated with it, partly because they are open to material and energy flows, but more profoundly because they structurally reflect their environment through the process of embodied cognition. Followers of the theory of autopoiesis emphasise that the structural embodiment of cognition is not a representation of the outside world, it is a 'bringing forth of a world' - one that is relevant to the cognitive organism. This term 'bringing forth' means realising (literally making real) those features of the environment that are responded to by the organism in the forming of its own structure. The abiotic world does not inherently have features, it only has particles located in space and time and flows of energy among them caused by forces acting among them. This continuous configuration of particles, flows and forces itself embodies information (by virtue only of it being a particular configuration, different from any other). This is where Bateson's famous quote comes from: it is only the differences (e.g. in particle locations) that make a difference to an organism that he defines as information: "information is a difference that makes a difference". What is meant by making a difference is that the organism perceives it and responds to it by adapting its internal structure, whilst maintaining its organisational closure. Maturana and Varela call this adaptation of internal structure 'cognition' and consider it a form of learning. The world is then the set of perceived differences that make a difference and it is this set of 'features' that constitute the world brought forth.
We can go a little further using our axiom that physical reality is an assembly of matter/energy configured (information) in space/time. When an organisationally closed system makes - for and of itself - a configuration that is a response to its perception of the external configuration, at a deeper level it is information correlating (pattern matching) with other information. That is, the cognition process is one of forming effective information (it would be worth consulting the definitions page to help understand this). In cognition, the information embodied by the closed system in its configuration self-organises to correlate with some pattern it responds to in the surrounding configuration of matter and energy.
Cognition - a wonderful insight
Those who have met autopoiesis before will know that a concept of cognition lies at the core of the theory. It still might seem strange to include what we normally regard as a mental process in the fundamental definition of life. On the face of it, this word meaning 'knowing' (having information and 'understanding' it) is out of place here; we are talking about a minimum living system after all. But Maturana and Varela's theory places cognition at the very centre of their definition of life.
They came to the realisation that "cognition is the process of living" after a breakthrough insight, which I reinterpret here in information terms. Recall that information cannot be other than embodied: all information is physical. Thus, cognition - gaining knowledge which is information - is the act of embodying information. Instead of the Cartesian duality of separate mind and body (physical structure and informational systems), we see that organisms (literally) incorporate both structure and information at the same time and by the same process.
Gaining information is the act of organising one's own physical components so as to embody the information. By "knowing something about the environment", what is now meant is "having a structure that encapsulates some information about the environment". Not only that, but according to the theory of autopoiesis, this structurally embodied information is not merely a representation of a feature of the external environment, it is a co-created part of it. Maturana and Varela say the embodiment "brings forth the environment", meaning it effectively creates it from the point of view of the organism who's structure embodies it. The reason is that the physical world does not have any features intrinsically. Features are not just a property of objects, they are a property of the interaction among objects: in this case they are an outcome of the cognition of one object by another. To say that environment and organism co-create reality seems over stating the causal power of cognition, until one recalls that as well as organisational structure, autopoiesis involves dynamic action. The action includes selection of metabolites and rejection of catabolites and this physical activity which alters the concentration of chemicals on either side of the organism's boundary does indeed create a new environment.
The relational definition of cognition - the two-way interaction between organism and its environment - is, of course, very close to the definition of semantic information: for data to have meaning, it must have an effect within a 'receiver' (observer) system - this effect being at least the creation of mutual information between the observed and the observer. The mutual information is the matching of pattern between one object and another. For an organism, this is equivalent to its adaptation to the environment: it orders itself in such a way as to respond functionally to a pattern in the environment by embodying something of that pattern. It thereby identifies and specifies the pattern: "bringing it forth" through the creation of mutual information. Such adaptation is an essential part of living (nothing lives unless it does this) and it amounts to embodiment of information in functional form. But what Maturana and Valera called 'cognition' was much more than merely reflecting a feature of the environment in its own structure. They meant the act of selecting the feature, indeed of transforming it from being a pattern to being functional information - i.e. 'bringing it forth'. Add to this that the functional result of the embodied information is that the organism changes relevant patterns in its local environment (e.g. by selectively internalising metabolites) and we see cognition as an embodied process that relates the organism and its environment in mutual co-creation.
This selective and active cognition is in fact what makes autopoiesis special to life, Without it, all we have is a system that creates and maintains itself and we could argue that, for example, the autocatalytic vesicles of Zepik et al. (2001) and Luisi's early experiments (detail below) can do that, but are not alive. Actually they can only do it in specially contrived circumstances where they have all that they need without the capacity for cognition (as we will see next).
We might (somewhat cheekily) paraphrase Descartes with "I embody features of my environment as part of self making, therefore I am".
More seriously, we might summarise Maturana and Varela's idea of cognition with the following three words:
Selection - Reflection - Action.
The first refers to active perception - the identification of salient patterns and creation of features out of these: this involves perception systems.
The second refers to the creation of mutual information with the features, embodying the reflected information in the structure of the autopoietic system.This involves structural assembly systems.
The third refers to the active process of manipulating the environment based on what is found in the features and this using the very structure that was created through reflecting them: as we shall see shortly, this involves actuator systems.
The three subsystems referred to above will be 'fleshed out' in the next major section. First a little bit more general thinking.
Autocatalytic Chemistry
Now we get less philosophical because autopoiesis is also a practical and real-life phenomenon. So let us next consider the way autopoietic systems might be implemented in physical practice.
The first attempt at testing the theory by making a minimally autopoietic system was a software simulation based on cellular automata. Of course this only serves to examine the principle of self-making, but it did work (Varela et al. 1974). The simulation has been superseded by the more sophisticated computer simulations of autocatalytic networks inspired by Stuart Kauffman's work. These were put into material practice by a series of chemistry studies, the leading light in the field being Pier Luigi Luisi (pictured below), who has made important contributions to our developing understanding of how life may have originated.
To be truly autopoietic, a system must be able to gather resources from the natural environment so as to utilise (make useful) these for the purpose of self-making (either repair or reproduction). Intriguingly, some quite simple chemical systems have been shown to exhibit this capability. For example, Zepik et al. (2001) experimented with fatty-acid ‘vesicles’ (self-contained droplets) and was able to demonstrate their reproduction and self-maintenance by autonomous homeostatic processes.

Bitbol and Luisi (2004) showed that autopoiesis and cognition are in fact separately necessary conditions for life, not inseparably linked as Maturana and Varela's original theory stated. It could be that this is a difference of opinion about what is meant by cognition, but in Bitbol and Luisi (2004), the authors are very careful to use exactly the (radical) definition of cognition implied by the original theory (summarised by my three words emphasised above). They illustrate their point using a simple chemical system of self-replicating autopoietic fatty acid vesicles (much the same as the self-maintaining ones in Zepic, 2001). In the diagram below, vesicles are formed from a surfactant S (essentially they are soapy bubbles in water). S-S is the (highly lipophilic (oily)) precursor of S (what S is made from) and it binds to the soapy membrane (the lipophilic bilayer). In this place, S-S is hydrolysed (water molecules added under thermodynamic control) to form the S molecules that compose the membrane. As this proceeds, the membrane surface area increases because there is more S in it and eventually the bubble gets unstable, wobbles and when two sides eventually touch, it bifurcates into two new vesicles: self-replication. As long as there is plenty of S-S in the environment, the more vesicles that are formed, the more S-S is bound onto the ever increasing total membrane surface area, so the process is autocatalytic. Since it all happens on the membrane which is a total enclosure (tegument) autonomously making itself, the system meets the requirements for autopoiesis: a) that it builds its own boundaries; b) that this process is only due to reactions taking place within the system and c) that it is performed entirely by reactions that are determined by the system itself (Varela's (2000) requirements).
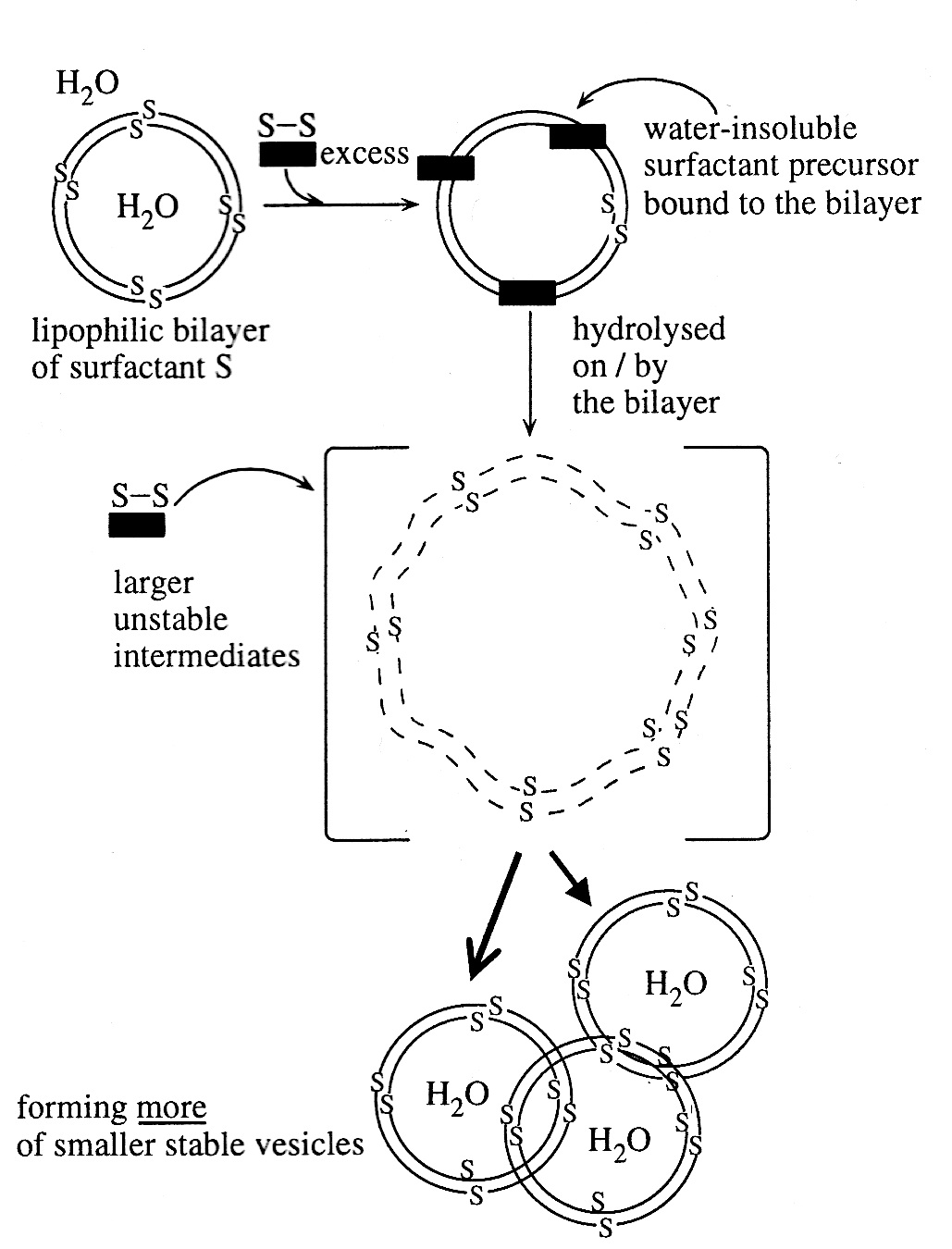
Figure 1 from Bitbol & Louisi (2004). Autopoietic 'bubbles'
It should already be obvious what is missing from this autopoietic, but non-living system. Self-replicating vesicles are not really meeting any of the three words (above) characterising Maturana and Varela's idea of cognition, and yet the system does match the definition of autopoiesis. Clearly autopoiesis and cognition are not synonymous. Bitbol and Luisi (2004) said this was because they are separately necessary for life. They also showed what cognition could be in the case of a minimal living system: it turned out to (literally) be metabolism.
Firstly, metabolism involves the selection of particular metabolites and the concentration of these within the enclosing membrane, leaving the concentration 'outside' lower. Conversely, it involves the ejection of catabolites with the opposite effect on the environment. Metabolism therefore entails selection, i.e. recognition (perception) of chemicals and action: moving them across the membrane against thermodynamic gradients. Metabolism, they say, always involves interaction with the environment. Obviously metabolism is the foundation of self-maintenance and self-construction; it is where the process of forming the chemicals and from there the interactions, that compose the living system, is to be found. What is found there is a chemical system that reflects the chemicals available in the environment. With this description of metabolism, we have all three of the words describing cognition.
Flexible and appropriate responses
Still, many would correctly point out that living systems are also quintessentially adaptive and flexible, in principle able to respond to any novel stimulus (that is not strong enough to kill them - a nod towards Nietzche*). Maintenance in a constant environment is a matter of homeostasis, in the face of a predictably changing environment it is adaptation, and in an unpredictable environment it is flexibility of response. Much has been made of the difference between homeostatic self consistency and responses to novelty. Like most biologists, Varela expressed the view that making novel responses to new features of the environment is a defining characteristic of life: organisms change their internal state in response to novel features without losing their identity (a tree may grow at a bizarre angle once exposed to a strong prevailing wind, but it is still the same tree; a bacterium may switch from aerobic respiration to fermentation - a radical change - when oxygen runs low, but it is still the same bacterium). Keeping the same identity despite internal changes is of course entirely due to maintaining organisational closure (see the autonomy page for explanation). However, it seems to me that the apparatus needed for homeostasis is essentially all that is needed for the flexibility that changing and unpredictable environments demand. So where Bitbol and Luisi (2004) distinguish between an "ordinary homeostatic metabolism" (with a limited range of responses) and an "open-ended metabolism of elements that are 'novel' in the strongest sense, since they require the domain of biology and cognition", I would say it is only a matter of scale. The 'structural plasticity' implied by the open-ended metabolism, with its facility for "unprecedented rearrangement of the chemical pathways" is still a homeostatic system having the 'goal' of maintaining the identity of the whole organism and a range of possible biochemical configurations to choose from, each better suited to a particular environment that can be recognised by the perception system of the cognitive whole. I will illustrate this in the next major section.
* Who is said to have claimed "any adversity which does not kill me will make me grow stronger" - a peculiarly apposite statement in the current context).
Putting it together - all life is one
From this work, it becomes clear that for a system to live, it must have at least the following three properties: autopoiesis, cognition (as described above) and an unbroken boundary to define its limits; this stops the ingredients of life from diffusing apart, rendering life's chemical reactions too thinly spread to work as a whole. This last element is a clear case of information embodied as living structure: the tegument or boundary selectively closes the living system in a thermodynamic sense, enabling a difference (reduced entropy) in the mixture of chemicals (and their energies) to persist either side of it (so reducing entropy). Living systems remain thermodynamically open in general terms, the closure is very specific and to some extent contingent on the internal state, which is in turn a response to the environment. What we see is a transformation from the necessity of cause and effect (characteristic of all non-living physical systems) into the contingency of stimulus and response, which is only seen in living systems. The tegument is not merely a boundary layer, it is packed with molecular-scale machines that sense and manipulate the environment both outside and in. Each one of those machines is a complicated pattern of matter created by the system within which it is functional. As it was with the vesicle membranes above, so it is a thousand fold with the living surface of the cell: made using the very processes which it performs.
In practice, all known living systems are cellular (though some biologists may include viruses). Significantly, the cell tegument has never been broken since the beginning of life - it has only been divided by repeated fission. Every cell in your body is not only a fission product (a daughter antecedent) of the first living cell, it is the most recent living instantiation of the first cells that had an identity and cognitively acted within their environment. Though none of the original material remains from approximately 3.7 billion years ago, today's cells are testement to the astonishing persistence of the autopoietic and adaptive design. You will note that multicellular organisms like trees and mammals are really just colonies of specialising cells that cooperate by communicating information and sharing materials. All of the cells are adaptations of the original design. In this way, the differentiation among organisms (such as you and I and the bacteria that live in us) is just an elaboration of differentiation among cell types that has been going on since the first common ancestor around 3.7 billion years ago.
All life from its beginning to the presently living, is unified as a the proliferation of cells which are the living embodiment of the autopoietic and actively cognitive system that keeps on self-reproducing and adapting. What has been replicated is material in only a rather trivial sense: living cells continually replace their material contents. What has been maintained and replicated throughout the whole history of life is the information content that organises this material into a structure that lives, by which we now mean it is information embodied in biochemistry and biological structure that maintains itself, using cognition and the information processing of self-control (homeostasis) and reproduction (information replication, which is a kind of communication) all made possible by the two unique features of life: autopoiesis and cognition.
We arrive at this: an autopoietic system is self-sustaining, homeostatic and autonomous, with a continous outer boundary, despite having a continuous flow of mass and energy through it. A living system is one of these which continually responds with active cognition to modify its internal arrangement (information) with the overall aim of maintaining its identity as a Kantian whole. Now let us briefly look at the physical components necessary to pull all that off.
The physical components of an autopoietic system
There are three conceptual components, though of course in a real autopoietic system, these are likely to be realised in multiple more or less complicated parts. The three necessary and sufficient components are 1) a boundary around the system; 2) a synthesis system for making the system from within and 3) at least one (probably a lot more) transducer-actuator system to enable controlled interaction with the external environment (mainly consisting of trans-membrane filters and pumps).
Multi-level homeostasis: perception-action systems
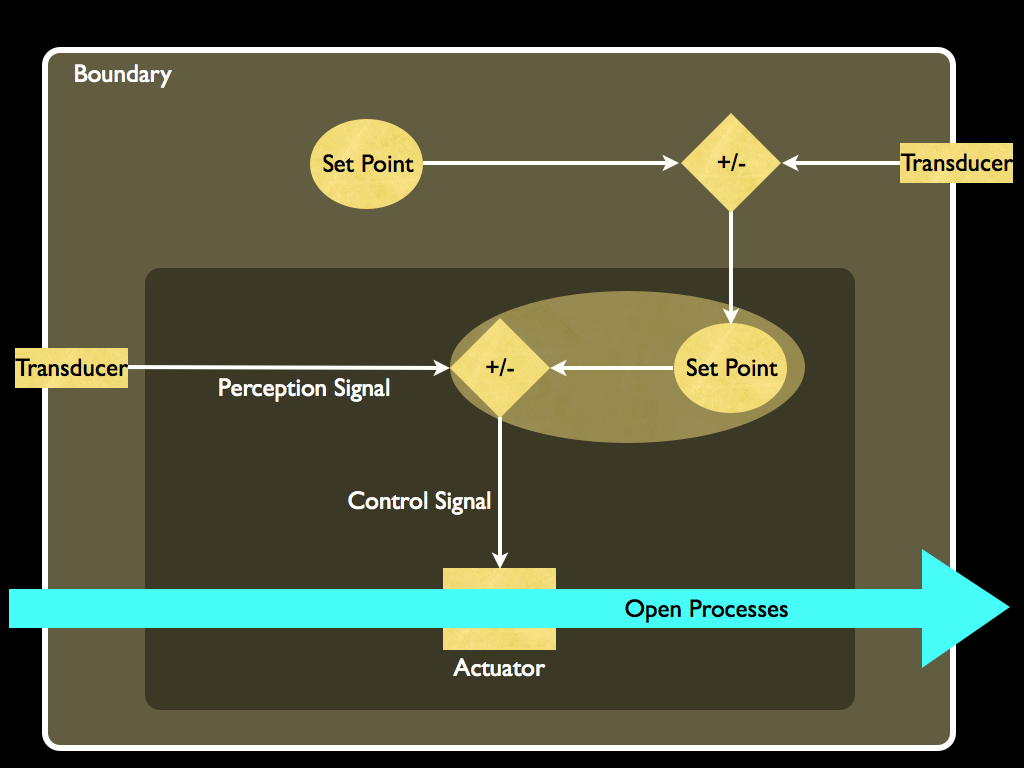
Part of this system that was only implicitly referred to by Maturana and Varela turns out to be very important in an information theoretic interpretation of autopoiesis. It is the set point-based homeostatic system. The control system of an autopoietic system are drawn together in the following diagram, which emphasises homeostasis.
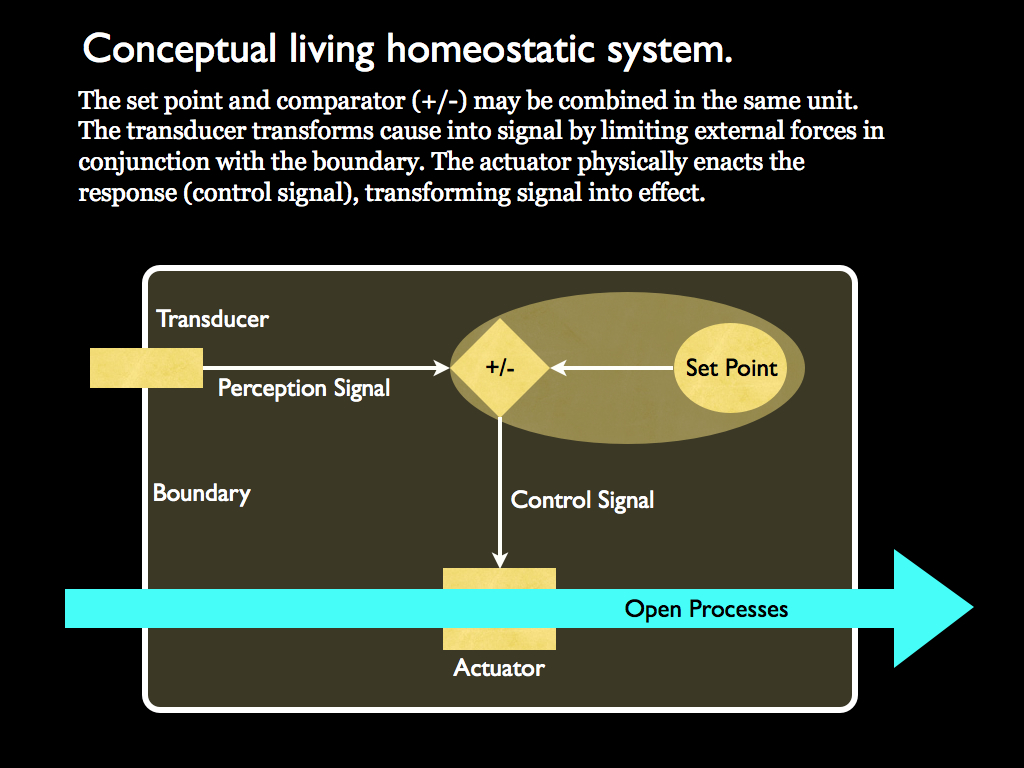
The system is contained within a physical boundary, into which are embedded transducers. It interacts with its environment as an open system: materials and energy can pass between inside and out across the boundary, but under control from within. The flow of materials is regulated by the actuators (also embedded in the boundary) and the behaviour of these is regulated by control signals that result from comparing an internally established and stored set point (information) with the signal produced by the transducers (perception).
The detail of the boundary and an example of an actuator is shown in the next diagram. All cells have at least an electrically polar sheet membrane acting as the boundary, usually it is a phospholipid bi-layer as illustrated. Within it are channels made of either single protein molecules or complexes of them. Many of these actively transport a particular, or class of molecules through and the rate at which they transport material can often be controlled (usually just 'open' or 'closed'). These channels take their control signal from an internal transducer which, in the case illustrated shows a protein molecule that changes shape (conformation) depending on the presence of an ion molecule and in so doing releases a messenger molecule that binds to the channel's 'gate': the switch that operates it (open or closed). There are lots of variations on this theme, but the essential ingredients are the same: a transducer detects something about the environment (either internal or external), generates a signal accordingly, this signal is responded to by a controllable channel in the boundary and by all this, the internal environment is regulated by selectively interacting with the external environment.
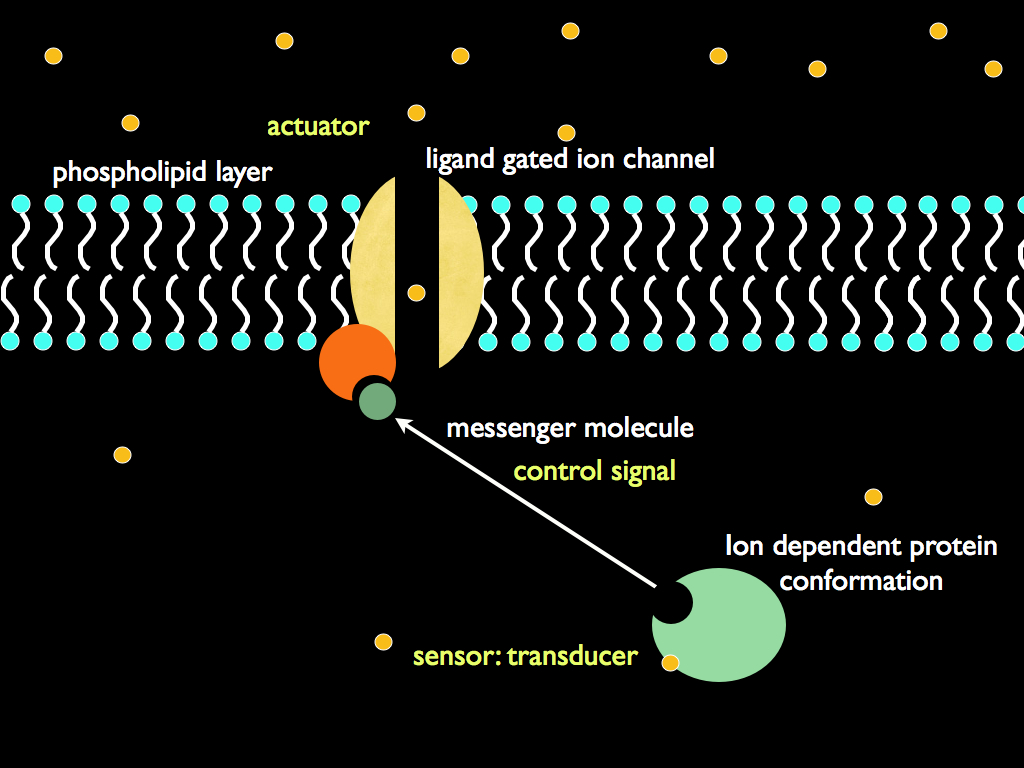
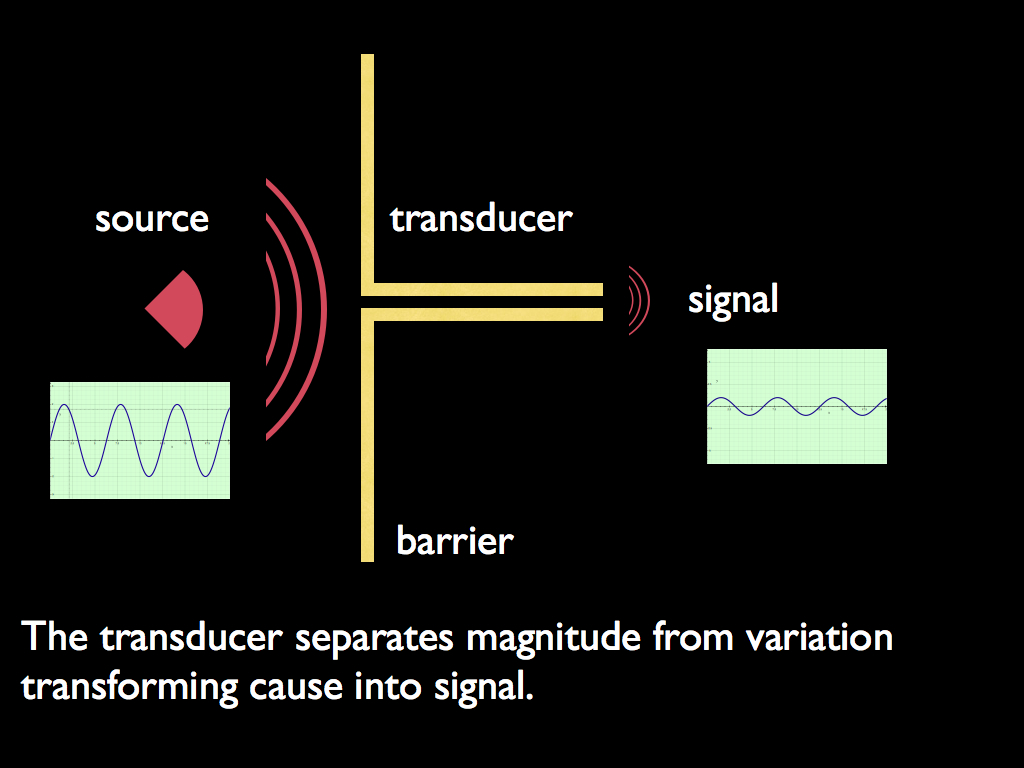
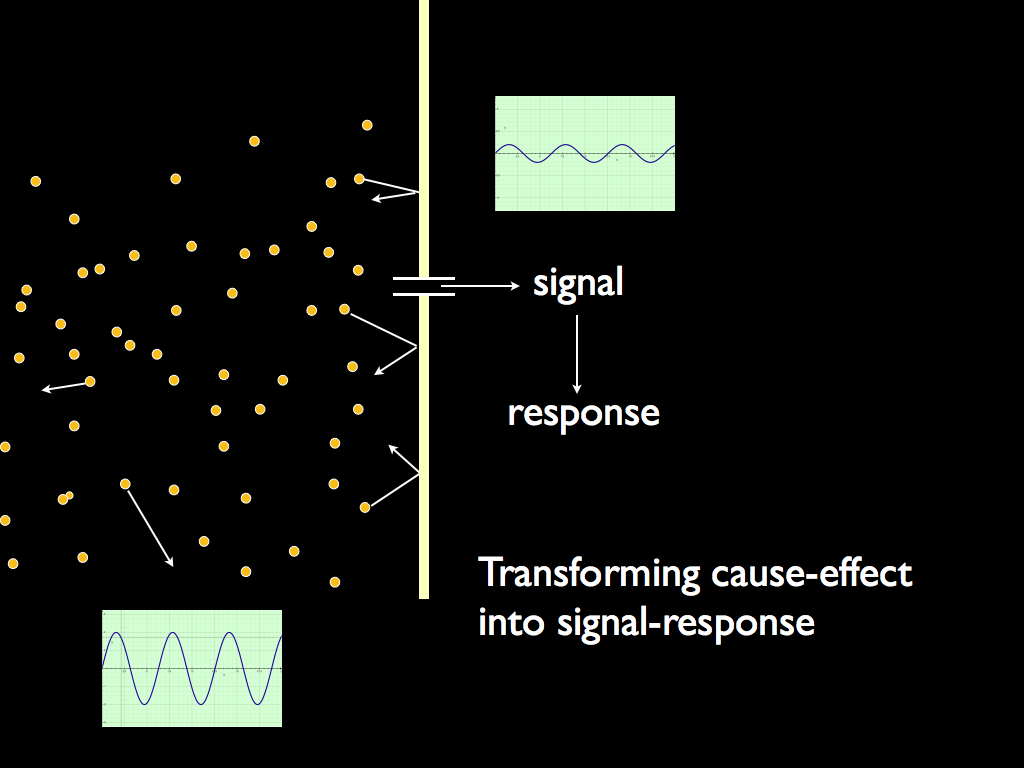
Transducers are the link between an organisationally closed system and its environment enabling the system to be autonomous. Without them, a closed system can be totally isolated - which is not much use - or else it would be at the causal 'mercy' of the environment, just like everything else. For this reason, they play a unique and easily overlooked but life-defining role. In conjunction with the boundary, they separate the variation in environmental quantities (e.g. molecule concentration or physical force) from the causal effect of the quantity. The variation is information, the causal effect is physical force. Essentially, transducers turn cause into signal and effect into response by translating from environmental variation into a specific signalling system within the cell: they are code translators. By being so, they break the inevitability of cause and effect. The internal signalling system responds to the external signals, buffered by the boundary and the transducer, but there is no inevitable connection between the external signal and the internal because the translation is defined (and can be controlled) entirely from within the cell. The special meaning of response here is that it is optional: the system can respond in one of a number of ways, or not at all. If it was not for the boundary (isolating the system from causal forces) and the transducers (translating information about the causal forces outside the boundary), then the system would either do nothing or would 'respond' in a way entirely determined by the causal force, just as any physical object would 'respond'. In other words, the transducers, in their boundary, confer one of two ingredients of autonomous action, which is the essential defining feature of a living system: they break causal dependence (the other ingredient is the 'private information' provided by the set-point that gives the system a 'personal' cause for its action (implemented as the choice of response it makes to the signal conveyed by its transducer).
The control system, consisting of transducer, set-point and its comparator and the actuator, with all the signals between them can be considered as a unit. A single cell can (and does) have many of these. Of great interest is the fact that a control unit can itself be controlled by another control unit (obviously implying vice-versa). This is illustrated in the diagram below which shows how an actuator can be regulated in a way that depends on the signal coming from another system and this consitutes an elementary form of decision making called a perception-action system. This active multi-valent homeostatic system is the foundation for what Bitbol and Luisi (2004) called an 'open-ended' metabolism: one that can produce "unprecedented rearrangement of the chemical pathways", yet maintain the identity of the whole organism.
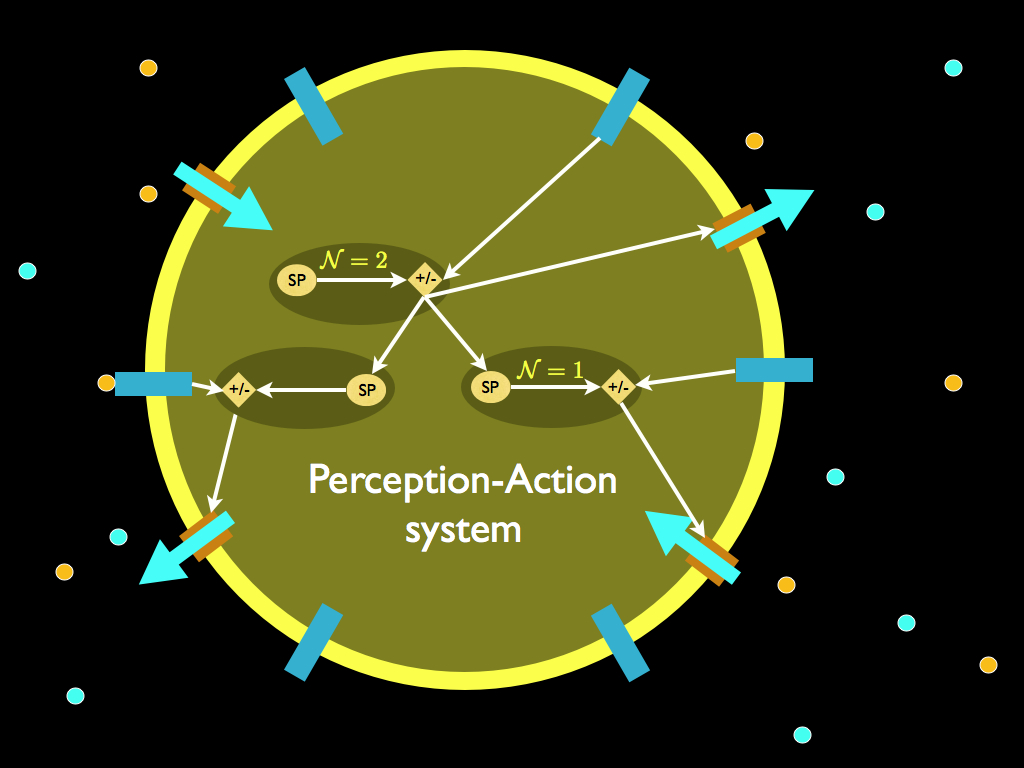
A more realistic looking cellular perception-action system is shown below. The calligraphic script N indicates the level of nested decision making: this is what was termed 'will-nestedness' in Farnsworth (2017). A variety of different transducers (blue bars) generate signals that are interpreted by multiple homeostatic subsystems who's actions are mediated through biochemical pathways that alter (internal) cell behaviour and simultaneously regulate (up or down) the chemical interactions between the cell and its environment via multiple actuators, e.g. gated ion channels (brown pores with flows passing through).
All that remains to add is a synthesis system, but that is easier said than done. In the next section we will see how the making of one's self "from within" requires " organisational invariance and metabolic closure" (Letelier et al. 2006), i.e. the preservation of a cybernetic structure during and as a consequence of its synthesis. Understanding this is the principle aim of Relational Biology and, more practically (compared to its very abstract mathematical description as (M,R)-System theory) by the autocatalytic set theory developed by Stuart Kauffman and formalised by Wim Hordijk and Mike Steel, though that is still lacking an account of structural invariance.
The synthesis system - making oneself from within
Uniting (M,R)-System theory and autocatalytic set theory in the context of autopoiesis is one of the core scientific programmes within the IFB project. Of course, a real (physical) synthesis system is composed of chemicals and their interactions and for autopoiesis, it is absolutely necessary for this to synthesise the enclosing boundary. To that effect, all be it in abstract chemistry, the model (below) by María Cárdenas and colleagues (Cárdenas et al., 2010) presents a mile-stone in understanding.
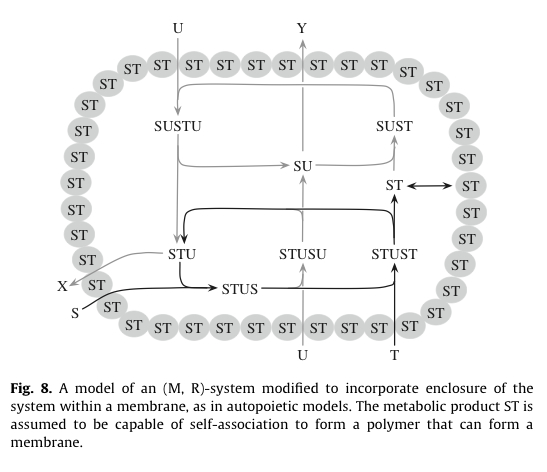
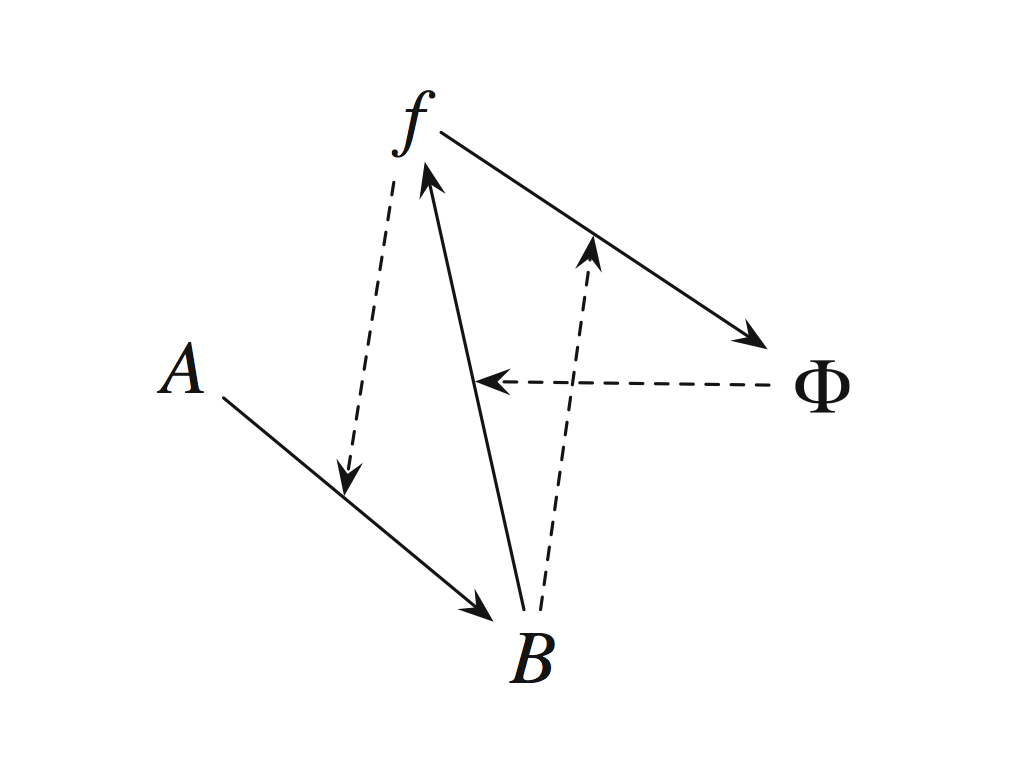 |
Figure 1.C. from Cárdenas et al., (2010) shows the interpretation of Rosen's (M,R)-system as an autocatalytic set (attributed to Goudsmit, 2007). Solid arrows indicate 'reactions', i.e. material transformations and dashed arrows indicate catalysis, interpreted as efficient cause. It is important to realise that the symbols here all represent much richer and more complicated ideas than mere chemical species, so the analogy with an autocatalytic set at this level of abstraction is rather misleading - see text. |
It is tempting to make a direct translation from Rosen's (M,R)-system to autocatalytic chemistry because catalysis has the character of causality: unaltered by its action, but enacting a change in a material system, where the 'material cause' of chemical reactants (the right chemicals) produces the products that are themselves catalysts, indirectly for their own production. The problem with that is that f and Phi and even A and B are not meant simply as chemicals in Rosen's very compact representation of "life itself". To be fair, neither Goudsmit (2007) nor Cárdenas et al., (2010) intended such a naive view when they drew the diagram reproduced above. This is made explicit and clear in their text. Taking substrates S, T and U (the food set in the language of Hordijk and Steel), Cárdenas et al., (2010) say that A is the (single element) set
{((S,T),(ST, U),(S,U))}, where ST means a molecular combination of S and T (and is an intermediate 'metabolite'), and Phi remains a function (not a molecule) which "assigns molecular identities to the enzymes [catalysts] involved in [the anabolic material transformations]":
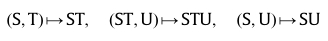 .
.The mapping arrow here denotes an operation of the form ST = f(S,T) in which the operator f is interpreted as efficient cause (though f is omitted in the relations above, copied verbatim from Cárdenas et al., (2010), if it were included it would be placed above the mapping arrow). Indeed, earlier in their article, they provide an autocatalytic set instantiation of the (M,R)-system in which f is identified with STU, but emphasise that this is just a (pedagogic) example: the 'real thing' is much more complicated of course. So, in Fig. 8 of Cárdenas et al., (2010) (above), S, T and U represent raw material (food) substraits and as they say in the caption, the molecule ST spontaneously forms a polymer with the structure of a membrane by self-assembly. This is clearly catalysed by the STU which performs the role of both Phi and beta in Rosen's analysis, so in the end these do appear as material in the Cárdenas et al., (2010) model: every element is embodied by chemicals. The black (not grey) arrows represent explicit chemical flows (metabolism).
This abstract formulation should be compared with the more biologically realistic (in relation to extant cellular life) produced by Jan Hofmeyr (2007) and redrawn below:
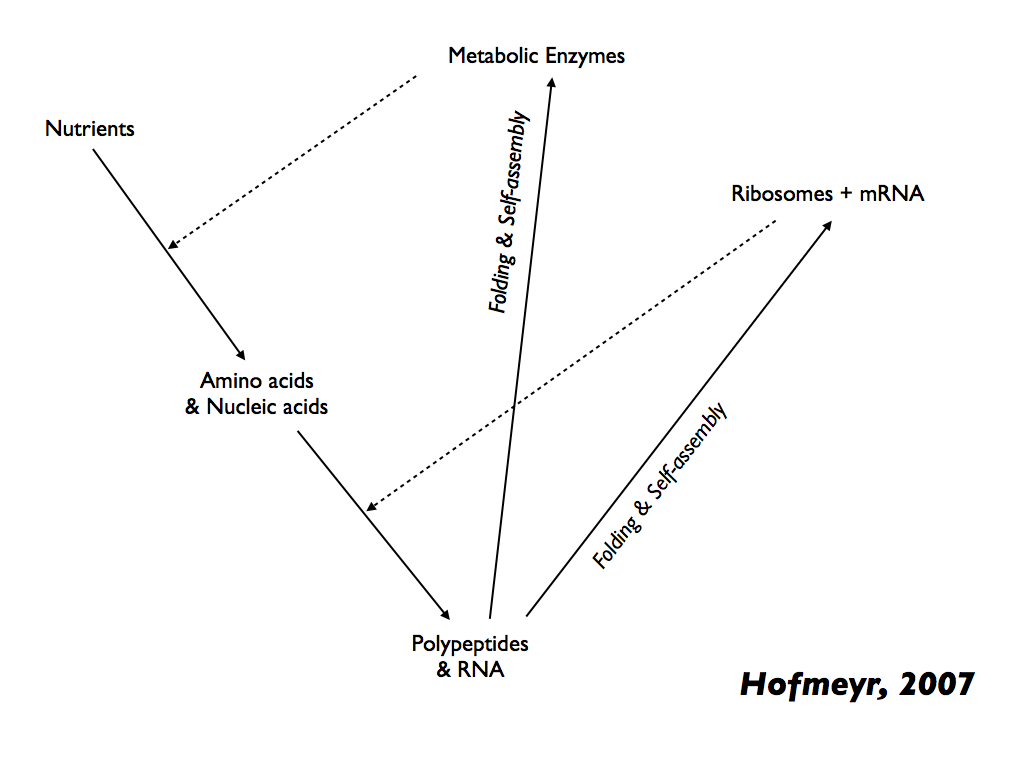
This arrangement looks strikingly similar to the (M,R)-System diagram shown in Cárdenas et al., (2010) (above) and deliberately so. Solid arrows show material cause and dashed arrows show catalysis, which is efficent cause. We now see, for the first time, proteins explicitly made from amino acids, themselves assembled, under catalysis by enzymes, from raw materials (material cause) with the efficient cause (like catalysis) of Ribosomes and mRNA (additional helper molecules assumed). Ribosomes and associated molecules are themselves assembled from raw materials in the same (conceptual) steps, so are the enzymes and therefore we have constructors constructing themselves. In fact, from the nutrient raw materials, you can follow two loops (one through enzymes and the other through Ribosomes) which combine to form closure to efficient causation and of course autopoiesis. Ten years later, Prof. Hofmeyr published a more detailed and biochemically precise diagram shown below (Hofmeyr 2017).
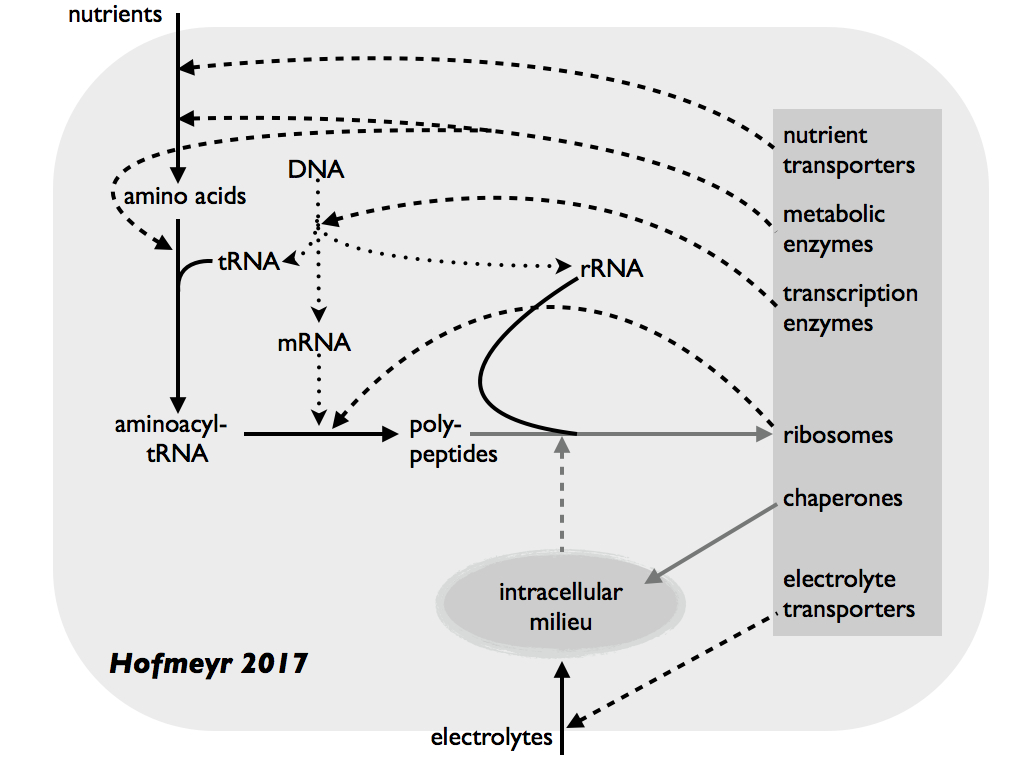
Now, dotted arrows add to the solid (material cause) and dashed (efficient cause) to indicate formal cause by sequence information (functional code); that is information which informs the efficient cause. Hofmeyr (2017) emphasises that, by providing an environment in which peptide folding (tertiary structure) leads to functional forms of proteins, especially enzymes and transporters (grey box), the intracellular milieu (especially including chaperone molecules) acts as an efficient cause in its own right. The grey arrows indicate supramolecular processes and what they depend on, e.g. the efficient cause of folding RNA and polypeptides into functional ribosomes. Again, one can follow the arrows to find a set of loops that collectively constitute an autopoietic system. [Note the versions of Hofmeyr's diagrams shown above were presented in Farnsworth, 2021].
References
Bitbol, M. and Luisi, P. (2004). Autopoiesis with or without cognition: defining life at its edge. Journal of the Royal Society Interface, 1(1):99–107.
Cárdenas, M.L., Letelier, J.C., Gutierrez, C., Cornish-Bowden, A., Soto-Andrade, J., 2010. Closure to efficient causation, computability and artificial life. J. Theor. Biol. 263, 79–92.
Farnsworth, K.D. (2017). Can a robot have free will? Entropy 19, 237.
http://dx.doi.org/10.3390/e19050237
Farnsworth, K.D.; Albantakis, L.; Caruso, T. (2017). Unifying concepts of biological function from molecules to ecosystems. Oikos, doi: 10.1111/oik.04171.
Farnsworth, K.D. (2021). An organisational systems-biology view of viruses explains why they are not alive. Biosystems 200: 104324.
Gatti, R.C.; Hordijk, W.; Kauffman, S. (2017). Biodiversity is autocatalytic. Ecol. Modell. 346, 70–76.
Goudsmit, A.L., 2007. Some reflections on Rosen’s conceptions of semantics and
finality. Chem. Biodivers. 4, 2427–2435.
Hofmeyr, J.H.S. (2007). Systems biology: philosophical foundations.. Elsevier, Amster- dam.. chapter The biochemical factory that autonomously fabricates itself: a systems biological view of the living cell. Systems biology: philosophical foundations. pp. pp 217–242.
Hofmeyr, J.H.S., 2017. Handbook of Anticipation: Theoretical and Applied Aspects of the use of Future in Decision Making. Springer. chapter Basic Biological Anticipation. 11, pp. 219–233.
Letelier, J. C., Soto-Andrade, J., Guíñez Abarzúa, F., Cornish- Bowden, A., and Cárdenas, M. L. (2006). Organizational invariance and metabolic closure: analysis in terms of (M,R) systems. J. Theor. Biol., 238:949–961.
Lu, N., Sanchez-Gorostiaga, A., Tikhonov, M. and Sanchez, A. (2018). Cohesiveness in microbial community coalescence. Available from BioRxiv at http://dx.doi.org/10.1101/282723
Maturana, H. and Varela, F. J. (1980). Autopoiesis and Cognition: the Realization of the Living. D. Reidel Publishing Company, Dordrecht, NL. English translation of original: De Maquinas y seres vivos. Universitaria Santiago.
Varela, F. J., Maturana, H., Uribe, R. (1974). Autopoiesis: the organisation of living systems, its characterisation and a model. BioSystems, 5:187-196.
Varela, F. J. (2000). El fenómeno vida. Santiago, Chile: Dolmen Esayo (cited in Bitbol and Luisi 2004).
Zepik, H., Blochliger, E., and Luisi, P. (2001). A chemical model of homeostasis. Angewandte Chemie-International Edition, 40(1):199–202.
Autopoiesis Resources on the Web
Tom Quick's PhD Pages Useful introduction, but not updated since 2003.
Encyclpaedia Autopoietica by Randall Whitaker - a massive compendium for those with a serious interest.