
What is Function?
You may think the word 'function', when used in a biological context, has a meaning that is self-evident and obvious. But on close inspection, it is often loaded with teleological and normative connotations which, as scientists, we are taught to avoid. For example, it seems reasonable to say ‘the function of the heart is to pump blood’, but what about ‘the function of predators is to regulate prey populations’? The latter is clearly not in the objective and impartial spirit of science. Often we could escape this problem by substituting the word ‘function’ with a phrase like ‘the effect of’, but doing that, we miss something important about the relationship between the cause and the effect, though it is hard to pin down what is missing. Here we are most concerned with what precisely is meant by ‘functional information’ - just what is ‘functional’ about this information?Function in Biology
In studying the precise meaning of the word function, used in the context of biology, Karen Neander (2011) has identified four branches of philosophical argument: a) Etiological; b) Systemic; c) Propensity; and d) Statistical. The essential insight transforming teleological and normative descriptions into more acceptably scientifically neutral and objective ones, is the recognition that evolution selects and shapes system components according to their positive contribution to organism fitness. Thus a function of X is identified as a feature of X that confers greater life-time reproductive success to the organism of which X is a part, than if that organism were without X. To this extent, we may say that X has the function and under the Etiological branch of argument, the function is what X was selected for. Systemic arguments more explicitly admit that this description is inseparable from the scientific observation and analysis of the system (or organism) into its apparently separate components, each identified by their contributions to the system’s biological fitness. The Systemic arguments run into controversy over the extent to which they retain a normative character: to what extent is ‘function’ - defined as a capacity of a system - a description of a virtuous character, so that its antithesis ‘dysfunction’ must be added for a complete description? One answer to this is a more explicitly adaptionist argument made by the Propensity branch, which defines ‘function’ as an adaptive trait of an organism. This of course requires explicit reference to the environment normally experienced by the organism, to which the trait may be an adaptation. However, since there may conceivably be ‘functions’ that are not organism traits and since traits are the outcomes of biological processes, rather than their causes, it seems a rather too narrow definition for our purposes. Another answer to the problem of malfunction or maladaption, is to say that only traits or capacities of a system which confer fitness are normally found: this leads to the Statistical branch of argument. With its own problems (see Neander 2011), it is not very clear how much further we get using a Statistical argument on function.Ideas for Functional Information
The Etiological proponent, Cummins (1975) proposed that function is an objective account of the contribution made by a system’s component to the ‘capacity’ of the whole system. Crucially, for Cummins, the capacity (meaning capability) of a system is explained in terms of the capacities of the components it contains, and how they are organised. This concept explicitly matches the understanding that functional information is to be found in the component parts and also the way they are organised into a whole. But it has been criticised, especially for its permitting what appear to be unintended consequences as functions (a frequently cited example being that dirt in a pipe may ‘function’ as a valve (Griffiths, 1993)). One of the solutions to this, at least for organisms, is to recognise that natural selection tends to eliminate potential functions of components if they do not contribute to the biological fitness of the system of which they are a part. This qualification was taken up by Neander (1991), by developing a biologically-based etiological theory (designed with physiology in mind). Whilst appealing, this cannot be used for all biological systems, such as ecological communities, for which evolution by natural selection has not been established, so to be general, we are forced back to the systemic theories of function. However, Darwin’s theory (and with it, neo-Darwinism) is a special case of a more general principle of selection in which the attribute of persistence is the superset of biological fitness (e.g. Kauffman, 1993). Thus we tentatively offer a definition of function that is systemic and in the spirit of established etiological definitions, but not reliant on Darwin’s theory. It is that any attribute A of a component C of a system S that causes an effect E such that S persists longer or in a wider range of conditions than without it, is a functional attribute of C. Then the functional information instantiated by C is that which establishes A, leading to the persistence in form of S, hence the persistence of information instantiated by S.An attribute A of a component C is itself a description of information embodied by the component. For example, the attribute may be the shape of a protein molecule. The molecule embodies information of form: the information instantiated by and fixing the position of each atom in the protein molecule. There may be several stable shapes to the molecule (conformations), each being a different packet of formative information and each summarised by an observer as an attribute: conformation 1 or 2 … etc.. Now if at least one of these conformations causes an effect E in the system S in which the protein takes part (for example catalysing a biochemical reaction) and if the effect E makes the persistence of the system more likely, then the conformation attribute can be said to summarise functional information. This functional information is of course the formative information that sets the conformation of the protein molecule. Now, consider an ecological community in which an organism lives and interacts. If the information embodied by that organism confers on it attributes which, in the course of living, cause effects in the wider community E, such that the community is more likely to persist, then we can say that the organism, and therefore its embodied information, is functional.
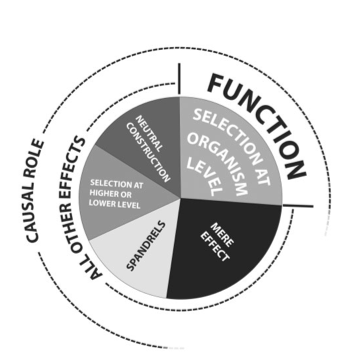
Ford Doolittle and colleagues (Doolittle et al 2014) used the diagram above to explain the different categories of effects that could be caused by genetic (or any other) information embodied in an organism. The most important distinction is between functions and 'all other effects': indicating that they were sure more than mere effect was needed to establish the existence of a function. Mere effects are incidental (e.g. the heart pumps blood (function), but also makes a noise (mere effect)). The next main division is between causal role -based function (CR) and selective effects -based function (SE). CR is not influenced by how the putative function arose, or why, it only refers to its potential action, as in Cummings function. SE on the other hand, critically depends on natural selection: it only admits as function those phenomena of the organism for which a historical (etiological) argument can be made to explain why the phenomenon arose and is conserved. Conventionally, biology only recognises natural selection on organisms (the assumed unit of selection), but this is clearly not complete. Natural selection is said to operate on genes (the selfish gene hypothesis), but its mechanism actually operates on the traits expressed by the genes (or anything else that gives rise to expression of traits). For his reason, SE functions based on selection at higher or lower than the organism level may be (at least) considered. A good example of higher level selection is that of symbiotic relationship, which gives rise to the sort of autocatalytic (so naturally selected) systems we see in ecology. The question these raise for function is: what is the higher ontological level of organisation to which this function contributes? The answer may be the ecosystem as a whole, though many biologists find that hard to swallow. In the diagram, two other CR candidates are admitted: spandrels are unintended side-effects, but more than mere effects because they do have identifiable roles in the operation of the system, even though that was not evidently the 'motivation' for their creation (often spandrels have negative effect, such as the propensity for rapidly dividing cells to turn cancerous, but positive effects often seed the start of natural selection: the swim bladder of fish probably allowed the development of lungs and the (presumably) thermo-regulatory feathers of dinosaurs turned out to be the key to flight). Finally, given continuous random change in genomes, it is quite possible that neutral (non-selected) coding of proteins, novel gene-regulation systems and ultimately traits could appear: this is often cited as a source of the large amount of so-called 'junk DNA' in most genomes. It is not at all clear yet what are truly junk (having no effect) and what are neutral, spandrel or higher / lower than organism level functional elements. Whatever the answer, it is clear that for function, there must be a potential effect and it must be enabled by information that is embodied in the biological system and conserved by it. Many biologists hold that it must also be a consequence of natural selection (whether or not operating strictly at the level of organism). For adherents to this website (IFB-followers), The CR-based definition of function is the most useful and has been formalised for all (including ecological) function using the concept of functional equivalence classes and hierarchical information structure (see below).
Relation to Entropy
Obviously these definitions suffer the same weakness in relation to dysfunction as any of the previously discussed philosophical proposals. It seems that we cannot avoid a sense of value judgment about the outcome of an effect, but at least when we define it as ‘making persistence of the system more likely’ , we do make the distinction between function and dysfunction on objective and scientifically neutral grounds. It is merely a matter of definition, we are not saying persistence is good and the collapse of a system is bad. Still, there is some room for taking sides, even in science. This is because the asymmetry between function and dysfunction, so defined, matches that of time (the direction of time being only one way), which may be derived from the second law of thermodynamics and the inevitable increase of entropy it predicts. In this context, functional information is that which leads to a local slowing (or even reversal) of entropy increase, whilst non-functional and dis-functional either have no effect, or accelerate the inexorable increase of entropy (noting carefully that this refers exclusively to the local system, for universally, entropy is always increasing). We may then say that functional information causes a local dip in entropy that has the effect of maintaining lower entropy locally, at least for a finite time.Rosen's Idea of Function
The taboo connotation of 'purpose' conveyed by the term function was very much noted by Robert Rosen - a major contributor to efforts to answer the question 'what is life?'. He says the term arose with the machine metaphor for nature and we cannot deny that. He further points out that its "overtones of end, purpose, finality and telos" have caused embarrassment to those who used it to get away from these very metaphysical ideas. Explaining 'purpose' away as merely a description of natural selection fails to fully cover the exposed metaphysics, as we saw in the paragraphs above.
Rosen offers an alternative that is related to the interventionist concept used in identifying causation. Imagine a system which left alone exhibits a certain set of behaviours. Then imagine excising a part of it to see what that part does in the context of the system. This act of intervention in fact creates a new system - the one that has not got the part removed. Comparing the behaviours of the original and this new system reveals the function of the part in the context of the system. Rosen identifies any part having a function (i.e. there was a difference in behaviour when it was removed) with a relation between part and whole. He calls such a part a component and identifies 'component' as the elemental object of relational modelling. His idea of intervening as an experimentalist might remove some brain part to discover its function by observing the resulting change in behaviour is clearly an example of the interventionist approach to causation and now informs the informationist approach of Integrated Information Theory (IIT) (see Bexendale and Mindt, 2018 for details). His explicit identification of function as a relation between part and whole (or more generally between ontologicial levels n and n+1) is encapsulated in our own definition of biological function (below).
More broadly, Rosen's relation theory is a key idea for his understanding of organisation and is the foundation of his relational theory of systems. That theory, though not explicitly mentioned, is the underpinning of all that follows here (it is laid out in Rosen's book "Life Itself" (1991) - which is essential reading).
A more general definition of Biological Function
In 2017, Keith, with Larissa Albantakis and the Italian ecologist Tancredi Caruso made an argument for a more general but precise definition of biological function (published in Oikos as "Unifying concepts of biological function from molecules to ecosystems"). We started with Cummins Function: : a process performed by a component that is necessary for performance of a higher level process by the system of
which it is part. We built on this, using ideas of functional equivalence classes (learned from Denis Noble and George Ellis), the concept of a Kantian whole defined by causal closure (learned from Stuart Kauffman and developed further in "Living is information processing: from molecules to global systems" and "Can a Robot Have Free Will?") and from the work on autocatalytic sets by Wim Hordijk and his colleagues.
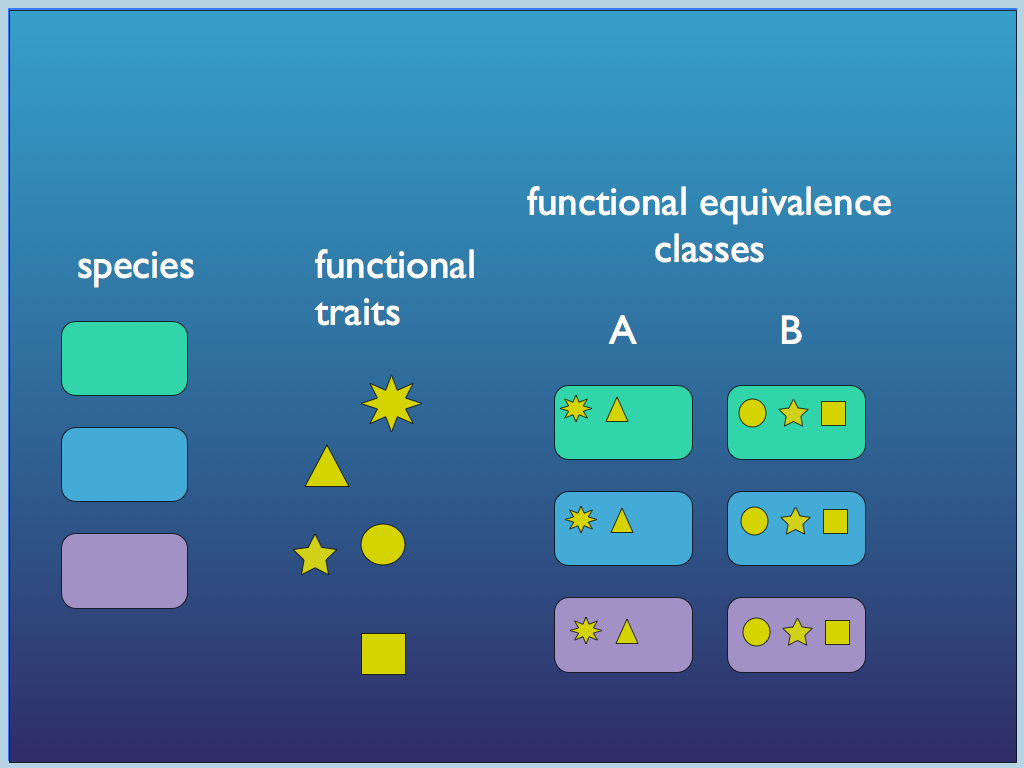
Our definition is independent of teleology (it does not distinguish between 'good' and 'bad' function) and also does not require natural selection at the level of organism, though it admits that for a function to be conserved, it must be naturally selected for at some level of organisation. Essentially our definition makes Cummings function more precise, particularly in specifying the connection between the sub-system that creates (or enables) a function and the system to which it confers a 'capacity'. It is a definition with roots in information theory and cybernetics, rather than neo-Darwinian notions of function.
The definition we came up with is this:
A biological function is a process enacted by a biological system A at emergent level n which influences one or more processes of a system B at level n+1, of which A is a component part.
An important advantage of this definition (apart form its precision and generality) is that it lays the ground for quantifying function. For that, we need to identify an objective function, i.e. the goal of the function (without meaning to be teleological: as "Can a Robot have Free Will" carefully explains). For organisms it is clear that Lifetime reproduction success (LRS is essentially Darwinian fitness) is the goal. At the higher level of communities of organisms, a good argument can be made for translating LRS into total biomass production. Using biomass production rate as an objective function, we illustrated the use of our definition of function to calculate the functioning of populations of different species of organism in a simple ecological community that contained primary producers, consumers, pollinators / seed dispersers and nitrogen recyclers. When we come to consider ecological structure, we will emphasise that all these are necessary parts of an ecosystem (food-web models are not sufficient). It is good to see that biomass production is something (a bit like money) that every other particular kind of functioning (e.g. seed dispersal) can be transformed (mathematically speaking, meaning something like using an exchange rate to transform between currencies) into. Biomass production turns out to be a kind of universal currency for biological function. Below the level of individual organisms, every part (e.g. the functions of your liver) contribute to your LRS via your bodily growth and reproduction of healthy offspring, so ultimately to your contribution towards biomass production. Sorry to put it that way!
An example with Bacteria
Bacteria have functions, lots of them, but not all bacteria have all the functions they need. This is because they are used to living together in communities. When people first saw this, they expected the bacterial 'species' to be competing with one another, but most of the interactions turned out to be almost the opposite and there is a good reason why they should be.
You might have heard of the 'Arts and Crafts Movement' - between the 1880s and the 1920s (more or less) groups of talented craft-artists quite often gathered in communes throughout Britain to make beautiful, humane and meaningful art (examples include William Morris). The communes consisted of people with huge expertise in, for example, woodwork, ceramics, metal work, painting and embroidery. They combined their skills to create beautiful objects that were often the product of multiple craft disciplines. The point about this is that they did not act alone and they did not try to everything - sharing and acting as a cooperative community, they became much more creative than they would have been alone. It takes thousands of hours practice to develop really good skills in any one craft discipline. No one person can achieve that in more than about two skills. If they had to turn their hand to others, they could probably get by, but the results would not be superb. When the skill is being learned, over those thousands of hours, information is being gathered and embodied in the minds of the artist. Time, more than anything, limits their ability to acquire skills to extremely high levels. For bacteria, with their very much simpler structures are limited by the total amount of information that can be embodied in them. A bacterium can try to do everything it needs to live alone, but none of its methods (functions) will be very sophisticated and probably therefore not all that effective. A bacterium species may be able to improve on this by dedicating more of its total information capacity to a few crucial functions, but then it would be jeopardising its ability to perform others. What if another species found an advantage in dumping some of its functions in order to concentrate on those that were missing from the first? Then the pair of them could live together, performing functions both of them need, but to a higher standard than either could achieve alone. This would be the evolution of cooperation. It would be motivated (and evolutionarily likely) if the fitness of both species increased as the level of specialisation among them increased. In many circumstances this does indeed seem to be the case.
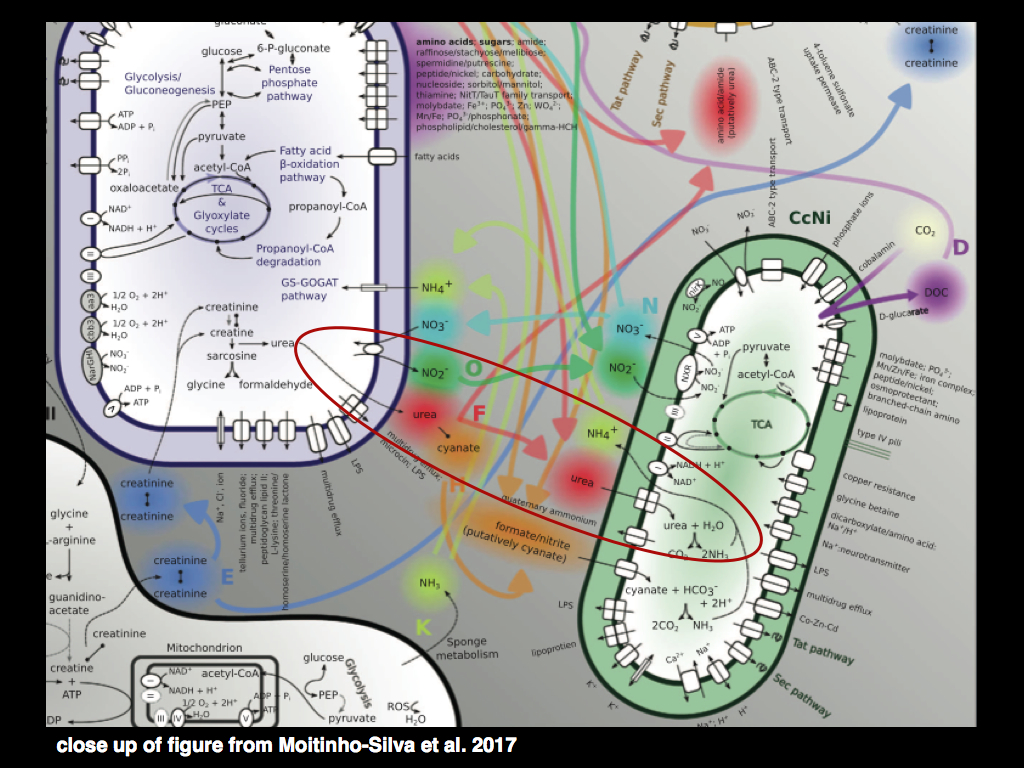
The diagram below (Fig 1 from Montinho-Silva et al, 2017) shows three species of bacteria, with a close-up of biochemical pathways. The purple one is breaking down creatine into urea, which it finds poisonous and has to get rid of. But the green bacterium has adapted to a world soaked in urea (pumped out of the purple bacterium) and it has an efficient process for breaking it down into ammonium and ammonia, getting a bit of energy in return. Meanwhile the purple bacterium likes to have some ammonium and ammonia, so it soaks up what the green bacterium has made... and where did that creatine come from in the first place? It is a waste product of metabolism ejected by the gray bacterium. Each of these processes is a function. Different bacteria have different functions and they tend to specialise in performing a limited set of functions really well, whenever they live in communities. The functions are distributed among the bacteria, the information required to instantiate the functions is distributed among the bacterial genomes. In an important sense, the species are little more than vessels for functions. What counts for this little community is the collection of well-honed functions and they way they all fit together to make an overall functioning whole. Species can come and go, what matters is the collection of functions.
References and further reading
Doolittle, W. F. et al. (2014). Distinguishing between “function” and “effect” in genome biology. Genome Biol. Evol. 6: 1234–1237.
Baxendale, M. and Mindt, G. Intervening on the Causal Exclusion Problem for Integrated Information Theory. Minds and Machines. http://doi.org/10.1007/s11023-018-9456-7
Cummins, R. (1975). Functional analysis. J. Philos., 72(20):741–765.
Ellis, G. (2008). On the nature of causation in complex systems. Trans. R. Soc. S. Afr. 63: 1–16.
Ellis, G. (2012). Top–down causation and emergence: some comments on mechanisms. Interface Focus 2: 126–140.
Farnsworth, K. D. et al. (2013). Living is information processing: from molecules to global systems. Acta Biotheor. 62: 203–222.
Farnsworth, K. D. et al. (2016). Living through downward causation: from molecules to ecosystems. In: Walker, S. I. et al. (eds), From matter to life: information and causality. Cambridge Univ. Press, pp. 303–333.
Farnsworth, K.D.; Albantakis, L.; Caruso, T. (2017). Unifying concepts of biological function from molecules to ecosystems. Oikos, doi: 10.1111/oik.04171.
Farnsworth, K.D. (2017). Can a Robot Have Free Will? Entropy. 19, 237; doi:10.3390/e19050237
Griffiths, P. E. (1993). Functional analysis and proper functions. British J. Philos. Sci., 44:409–422.
Hordijk, W. and Steel, M. (2004). Detecting autocatalytic, self sustaining sets in chemical reaction systems. – J. Theor. Biol. 227: 451–461.
Jax, K. (2005). Function and “functioning” in ecology: what does it mean? Oikos 111: 641–648.
Kauffman, S. A. (1993). Origins of Order: Self-Organization and Selection in Evolution. Oxford University Press, Oxford, UK.
Kauffman, S. A. (1986). Autocatalytic sets of proteins. J. Theor. Biol. 119: 1–24.
Moitinho-Silva, L., Díaz-Vives, C., Batani, G., Esteves, A.I.S. Jahn, M.T. and Thomas, T. (2017). Integrated metabolism in sponge-microbe symbiosis revealed by the genome-centred metatranscriptomics. ISME Journal. 11. 1651. (www.ncbi.nim.nih.gov/pmc/articles/PMC5520145)
Neander, K. (1991). Functions as selected effects: A conceptual analysts defense. Philos. Sci., 58(2):168–184.
Neander, K. (2011). Routledge Encyclopedia of Philosophy (Online). Routledge.
Noble, D. (2012). A theory of biological relativity: no privileged level of causation. Interface Focus 2: 55–64.
Rosen, R. (1991) Life Itself. A comprehensive inquiry into the nature, origin and fabrication of life. Columbia University Press.