
Complexity of Life
Critical steps in forming the complexity
of modern life
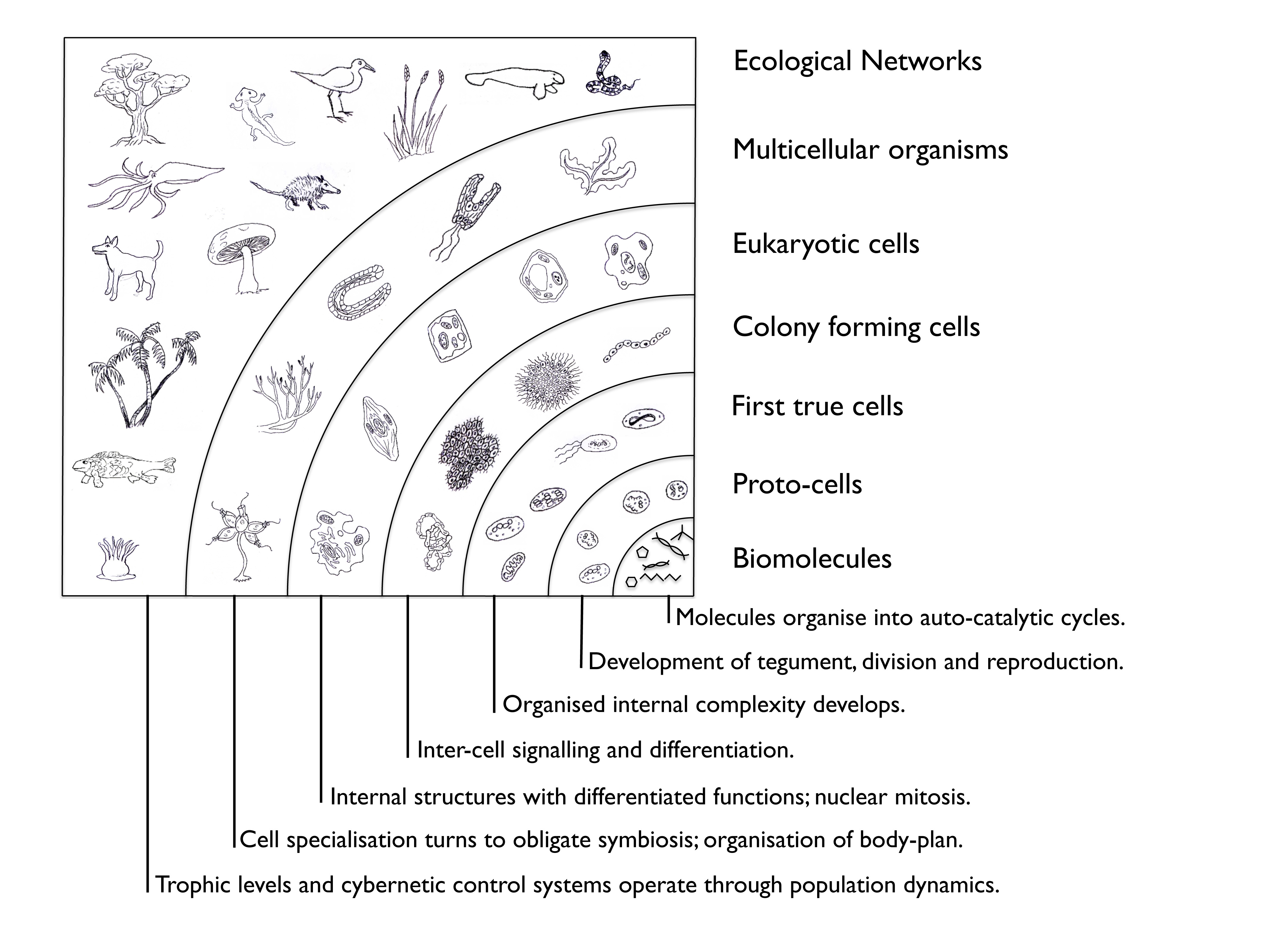
Diagram from Farnsworth et al. 2012; Creatures drawn by John Nelson.
Similar diagrams to the one above, are sometimes used to illustrate major developments in evolution, but the intention here is rather different. The diagram catalogues major steps in emergence: the information-dependent property of complex systems that creates new layers of organisation.
This diagram above is used to explain a sequence of steps needed to build up the complexity of life on earth as we find it today. It is important to realise, though, that the kind of life produced at each stage does not replace the previous stage, but coexists with it (and there are typically far fewer individuals of larger size with each higher level of complexity). Proto-cells are the only exception to this: it seems they have been thoroughly out-competed by the far more efficient true-cells that are the basis of life as we know it. The way true cells (autopoietic enclosed complexes of spatially and temporally ordered biochemical interactions) work more efficiently than any proto-cell is still not well known, but it is the essence of life as we define it and it is the reason we are all here.
As well as their internal organisational structure, some of the attributes of ecological networks have existed as long as these true cells. Certainly competition for finite resources among individuals has been a feature of (and a fundamental driving force behind) the development of organisms. Specialisation of functions also originates from the earliest proto-life stage. This enabled the ecological phenomenon of specialist classes of organisms acting collectively as a chemical processing chain, in which the products of one cell-type become the resources for another: enacting a kind of symbiosis. This phenomenon is still today very much a part of microbial community organisation (for example the human gut fauna is interconnected by multiple processing chains that break down different molecule types, with each organism class contributing a different set of functional capacities). Processing chains are likely to have been an important pre-cursor to the forming of colonies and eventual development of multi-cellular organisms.
The point at which colonies or networks of processing chains first coalesced into unified organisms is still not clear and it has never been reproduced in an experiment. All we have had to go on is the, difficult to interpret, fossil record and mathematical models. These models represent hypotheses about how the fitness (lifetime reproductive success) changes as independent co-operators first become obligatory partners and then give up their independence to the collective, becoming part of a multicellular organism with its own autonomy. The models reveal that the main impediment to that coalescence is the advantage that can be gained from ‘cheating’ within a co-operative network. Soon we may have genetic interpretations that show when, and in what organisms, the genes enabling multicellularity to emerge were first expressed. From the mathematical modelling, it seems that genes which suppress individual fitness optimisation, preventing cheating, are a good candidate for the search. These genes are the very ones that fail when we (and any other multicellular organism) gets cancer. Indeed there is a close link between the fundamental causes of cancer and the evolution of multicellulaity - it gives an urgently practical motive for studying this topic.
The formation of Eukaryotic cells represents a very special case of the coalescence process, as bacteria either invaded or were absorbed into the body of a single cell to become obligatory symbiotes. Mitochondria and chloroplasts are the best known examples of it. This process is called endosymbiosis and is an idea first proposed by Lynn Margulis: it was very controversial to begin with, but now widely accepted. The result has been the development of eukariotic cells: the ones from which all plants, fungi and animals are made.
Eukariotic cells tend to be much larger than their prokaryotic (bacteria and archea) cousins and some of them turned to a new method for obtaining their energy and material resources: they became predators. This heterotrophy (as opposed to the homotrophy of cells that make their own resources) is a rather specialised form of parasitism, in which the host is completely consumed in a relatively short period, dismantled within the body of the predator and used as raw materials and energy for its autopoiesis. The predator-prey relation was added to competition and symbiosis, along with more general parasitism, to make up the 'big four' ecological interaction types. Ecological communities are networks of these interactions among organisms of every kind. The extent to which ecological communities are organised (in the sense of emergent systems) is still quite unclear. Finding the answer to that is one of the research fronts in which we are involved.