

The Ma of Ecology:
organisational power in the empty spaces
A brief illustrated article by Keith Farnsworth, based on a lecture given at the University of Maine, 2018.
(best seen in full-screen mode)
(06-01-2020 I have a little more work to do on some of the details, including references).
(best seen in full-screen mode)
(06-01-2020 I have a little more work to do on some of the details, including references).
I believe that the organisation of
interactions among organisms in an ecosystem is neglected by modern
ecological science (especially among non-trophic relationships). If I
am right, we are misssing the forest for the trees and missing the
essential character of ecological systems that makes them work and, for
example explains the biodiversity-function relationship. I explain that this neglected character is pure information and potentially it has formal causal power and indeed acts as a transcendent complex
to give an ecosystem function in its own right. In other words, that an
ecological community is not just an assembly of speceis that happen to
get on with each other in a more or less stable way, but is in fact a
real thing that selects its member species for their functional traits. It is only a hypothesis at present (see PhD projects for a chance to proove it).

Hasegawa Tohaku - Pine Trees (Shōrin-zu byōbu) - right hand screen
'Ma' is a Japanese word which has no direct translation into English. It has strong cultural and mystical meaning.
It is said that the essence of a pot is the space it contains; the essence of a room is the space it encloses: space completes the form and gives it meaning.
Japanese culture names this ‘Ma’: the empty space that gives things meaning. It is exemplified with the typical economy of Japanese art in the beautiful screen printing shown above.
My question is this: can we apply the principle of Ma to better understand what an ecosystem is? Of course, the answer is going to be yes. Let us see why.
Structure
First, rooms and pots, etc. are physical objects that are what they are because of the specific placement of material in space and time: they are geometric structures of material substance.
When it comes to living nature, ecologists typically survey a natural community, count the species and the numbers of organisms of each and then call this ''community structure'' (see the biodiversity indices page).
A catalogue of species present and their relative abundances is not what most people would call structure.

This is more like it, though it is a human artifact, the Forth Bridge is an ordered set of material components in which the order (placement of each in relation to the others) creates a whole. It is an example of the general class of engineering structures.
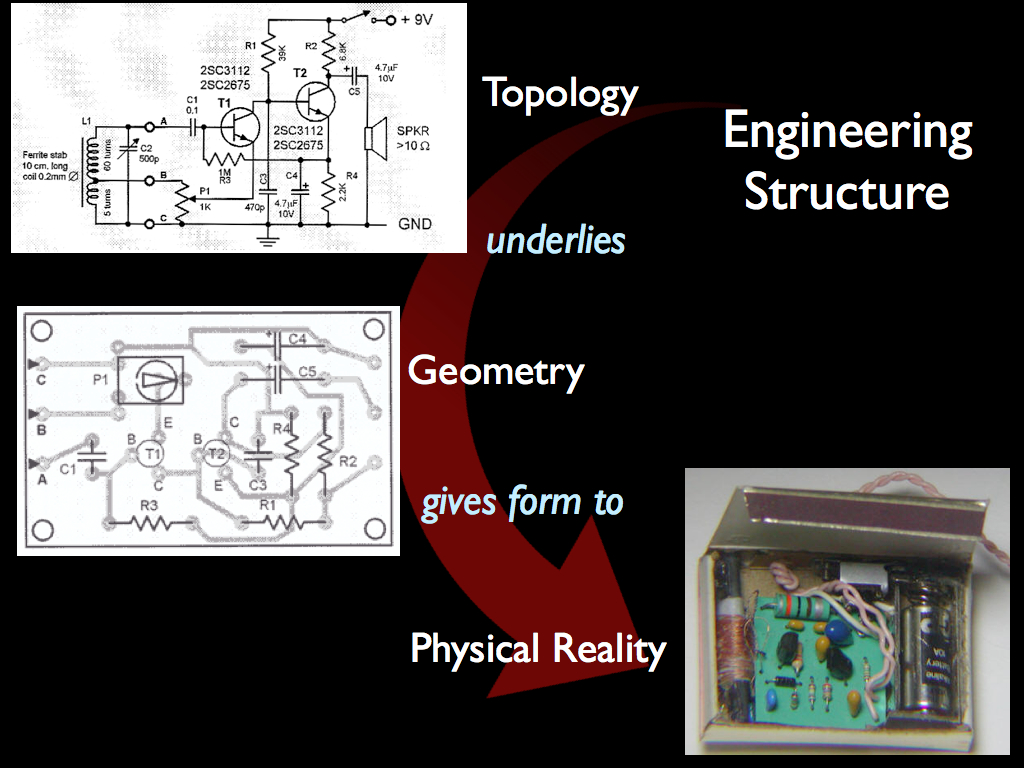
All members of this class share the following:
a) an underlying topology determines which components are connected to which and by what connecting points (be they connectors, nuts and bolts or points for welding etc.);
b) a geometry in which the topology is expressed in physical space;
c) a physical form, which instantiates the design using physical components arranged and interconnected in the real geometry. In the example illustrated above, a simple radio receiver is first specified as a topology connecting specific functional components in a particular way, this is then extended in space using printed circuit board tracks to make material the (electrical) connections and then the assembly, including the physical components, completes the structure. Please note that at every one of the three stages, organising (constraining) information is added to specify a particular configuration of parts, not any configuration, just this one will work as a radio. Is it possible that the same can be said for an ecological community?
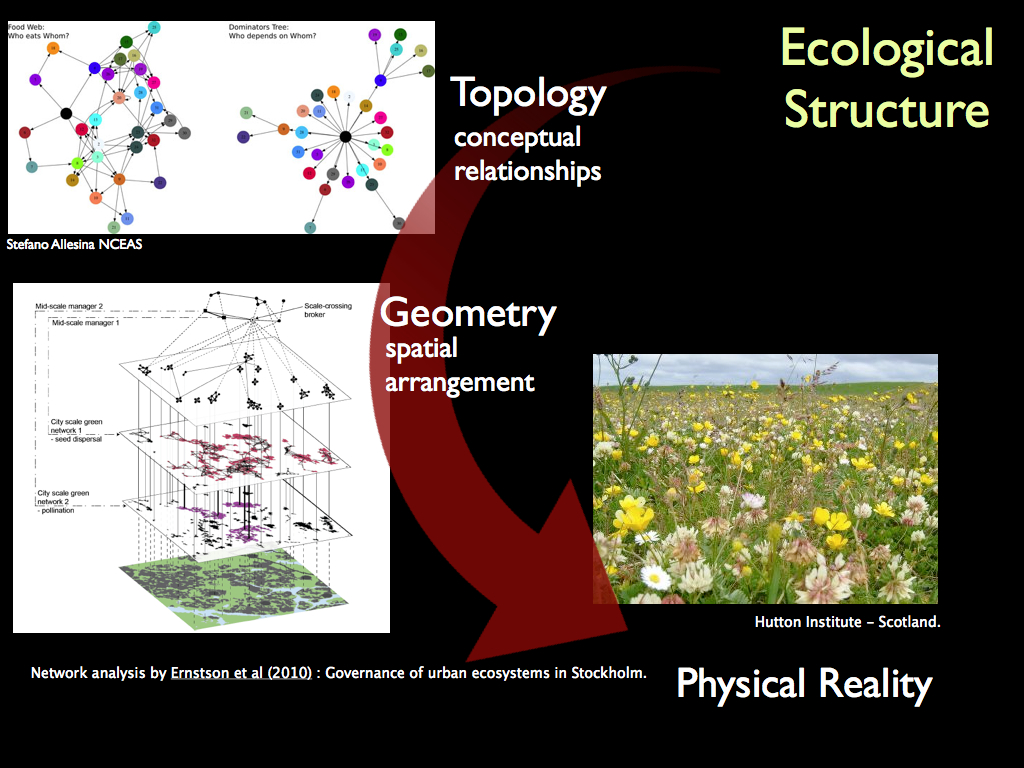
We can at least conceive of a model of such using the same principles, as this example (above) from Ernstson et al (2010) shows.
In general terms, the idea is that the information specifying the links of interaction among the component organisms constitutes the information component of the structure of the community. The other component is its physical material, just as in the Forth Bridge.
Embodied Information and Constraint
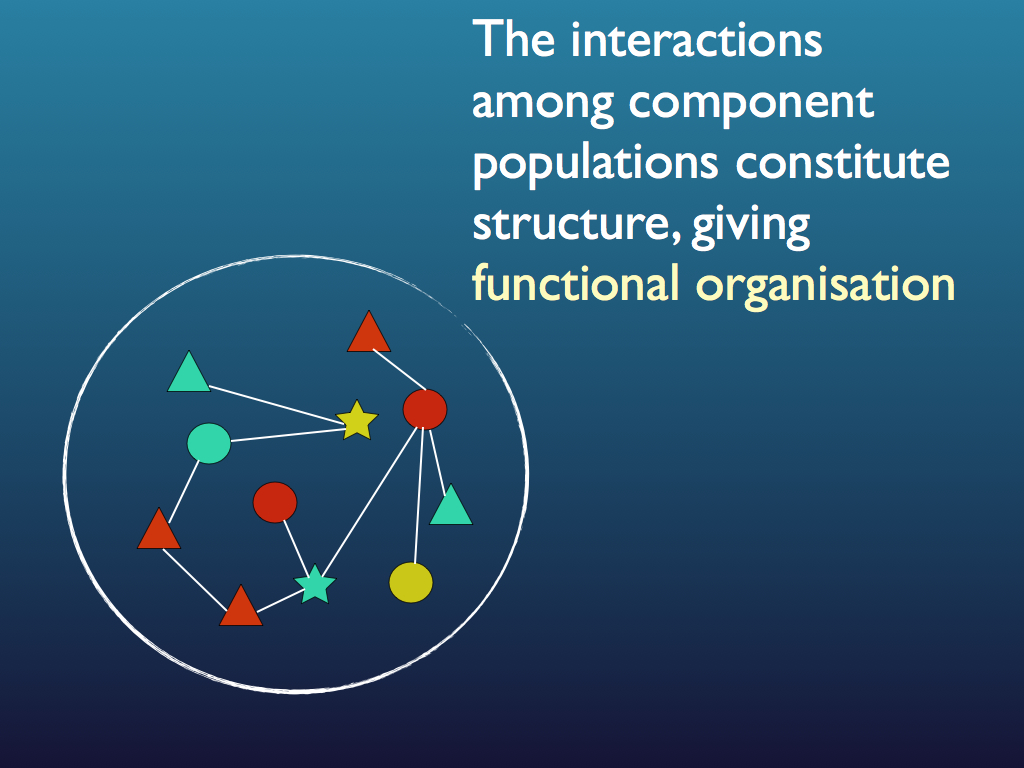
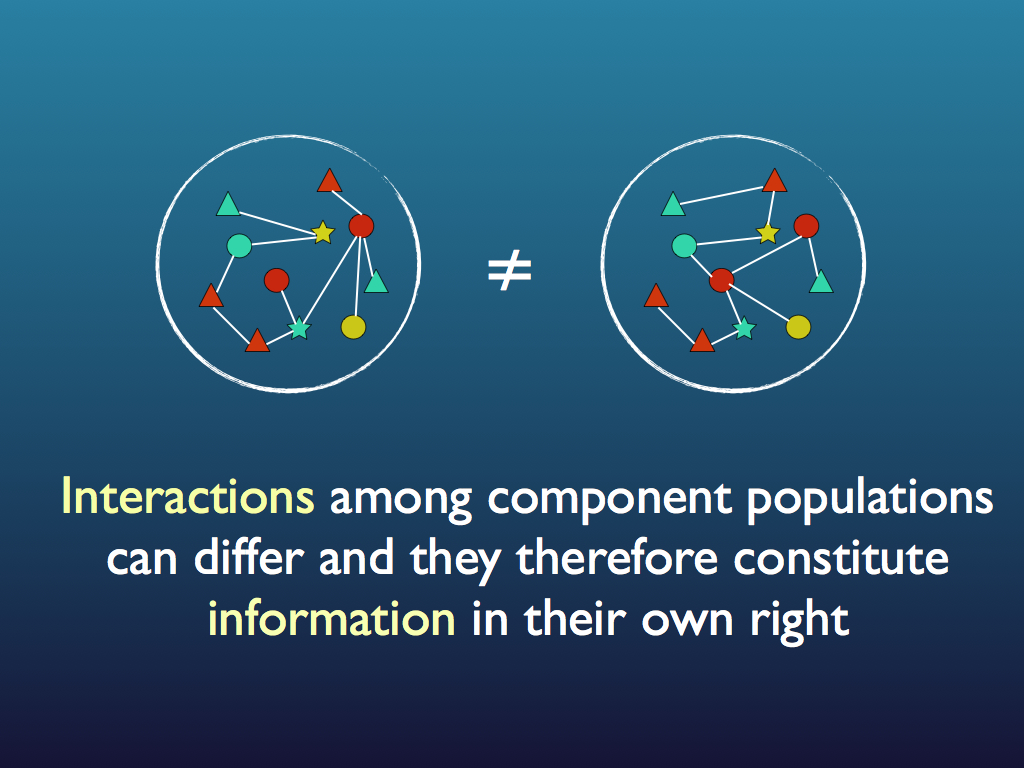
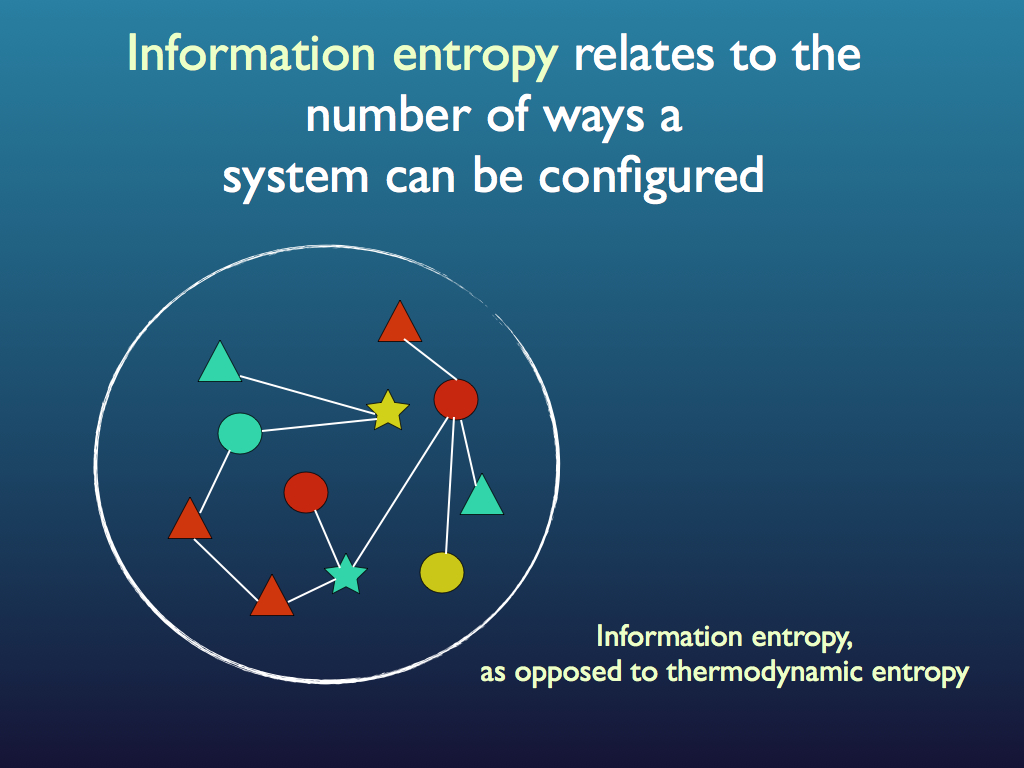
The set of interactions is information because it is specific to the community: recall that among the component we can have a large number of possible configurations of links and specifying just one from among all these is equivalent to a large reduction in the system's information entropy, which is equivalent to it gaining a large amount of information (see diagrams below and see the entropy page for a fuller explanation). Note that usually the precise configuration of links and components is not important to the community as a whole and in fact changes through time in a way that may as well be random. This is redundancy that is allowed because it is the functional relationships that matter to the community, not every detail of how they are created. Indeed, as we shall see shortly, the specific identity of organisms and even of species is not necessarily important to the community because it is only the functions they perform that matters at the community level. As such, the information is less than that of any exact specification of all interactions at a fixed point in time. The components, with their functions (based on functional traits) form a functional equivalence class for the community. This information is embodied at the system level as the set of interactions and it is functional because the system as a whole gains capacities from it, not least of which is the ability to self regulate and self-repair (summarised phenomenologically as 'stability' and 'robustness').

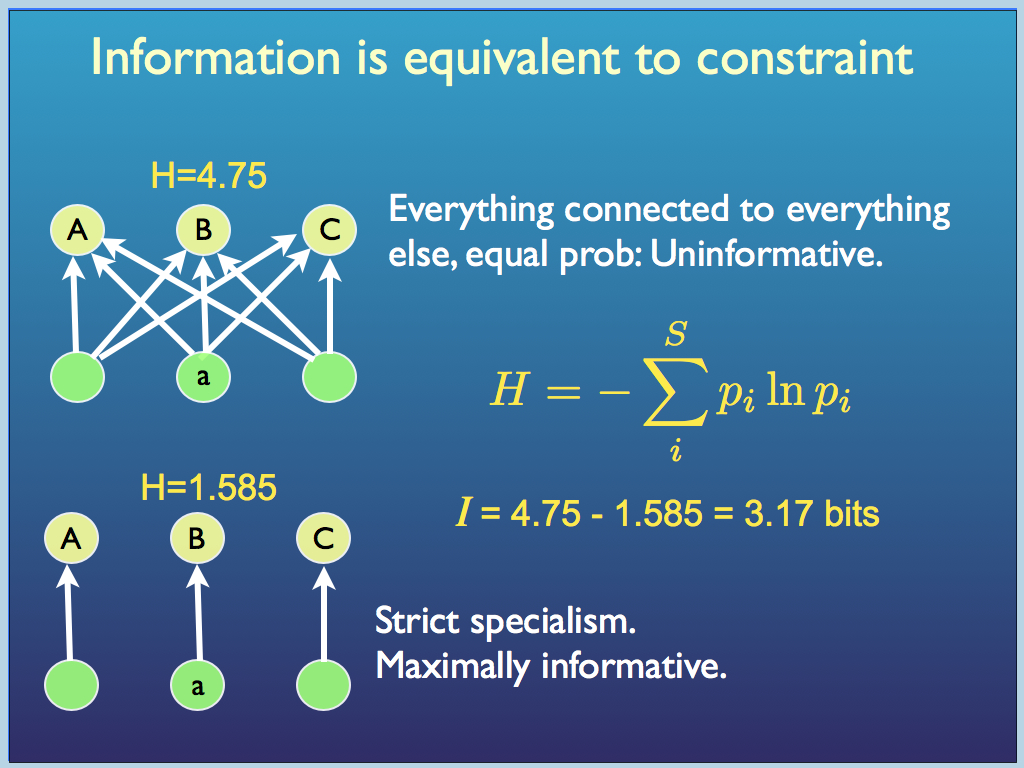
Taking the specific and very simple example (below), we can look at the difference between a network of interacting species that is a) simply a uniform interconnection (as for example in a community of generalist (opportunist) predators and prey, where the only 'rule' is that if a predator of any species meets a prey of any species, then the outcome is not good for the prey) and b) a community of specialists, in which each predator fastidiously only chooses to eat a specific prey species, leaving all the others. The generalist community has a network that is uninformative. We calculate its entropy using Shannon's formular, in which the p values are the probrabilities that a connection (predation incident) is made. In the uninformative community all the ps equal 1 and we get a system entropy of 4.75. But in the community of specialists, most of the ps are zero and the system entropy is the much smaller 1.585. That means that this specialist community has embodied information at the system level, an amount equal to the entropy of the most uninformative community possible, minus its own entropy. That works out as 3.17 bits of information in this little example.
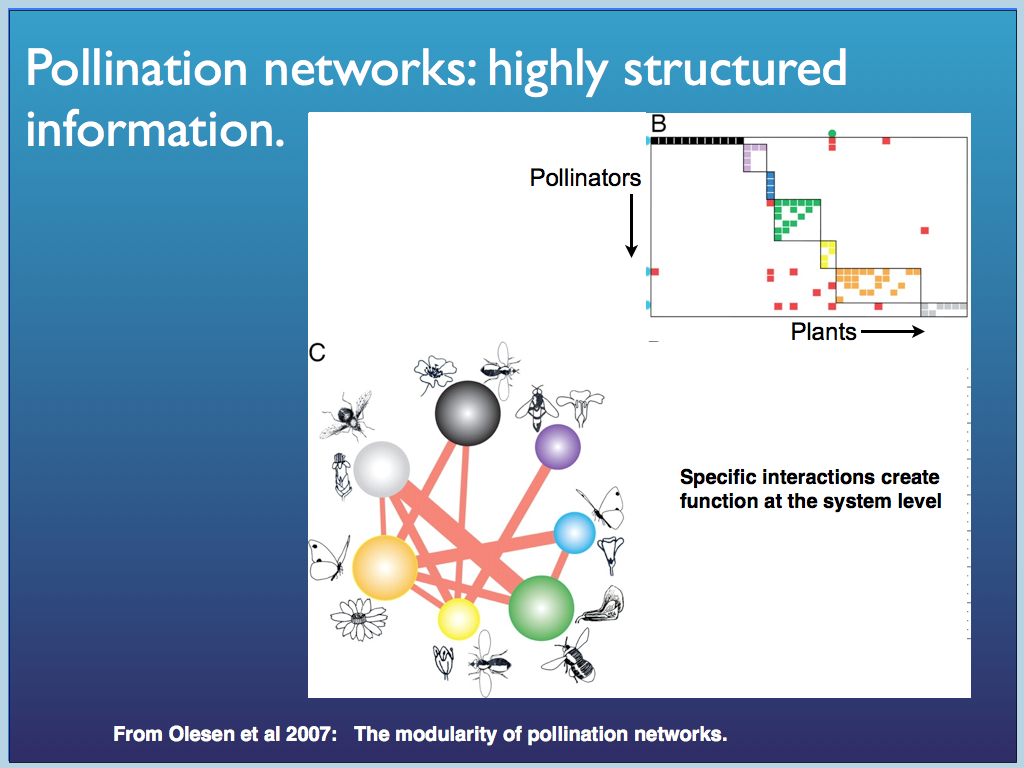
Now here is a real life example: the symbiosis network of interactions between flowering plants and their pollinators. I did not work out the entropy of this system, but given the data, we could do that. The point emphasised below is that the network is very far from uniform, uninformative. There are strong associations and very weak or near p=0 ones and this results in a distinct patter of interactions, which even contains a higher order structure, as shown in the graph of pollinator species against plant species (coloured squares showing strong interactions). This higher level structuring is revealed as clusters coinciding with the colours.
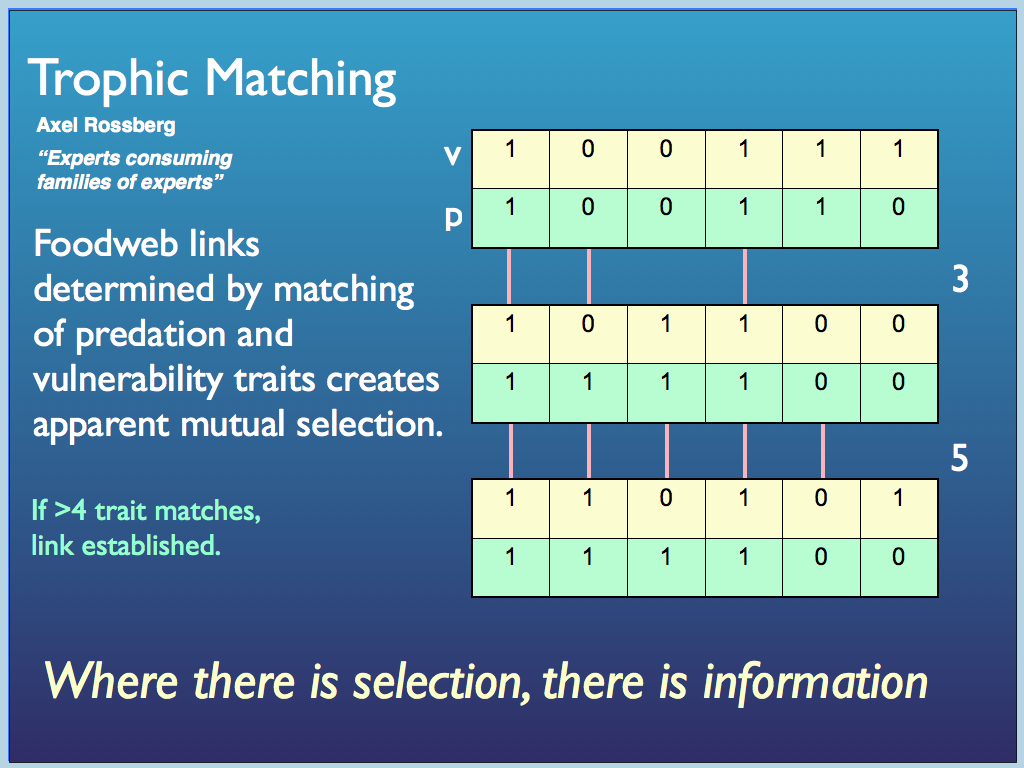
By far the majority of ecological networks studied professionally are trophic networks - foodwebs. My former colleauge Axel Rossberg has studied the effect and importance of non-random preferences of predators for particular species, building realistic models based on the idea of trophic trait matching. The idea is that a predator (a) is more likely to take prey (b) when the predator has traits that complement those of a prey. Taking a very obvious example, they both live in the same sort of habitat.
So, we may reasonably think there is a general pattern here.
The information caused by all the particular selections making up the non-random network of interactions is embodied at the community (system) level and is responsible for functions that arise from that level. We should recall what is really meant by function here. It is a capacity of an organisational level to contribute to the capacity of the next higher level. In Farnsworth et al. 2017,
we explain what this means at the level of ecological community.

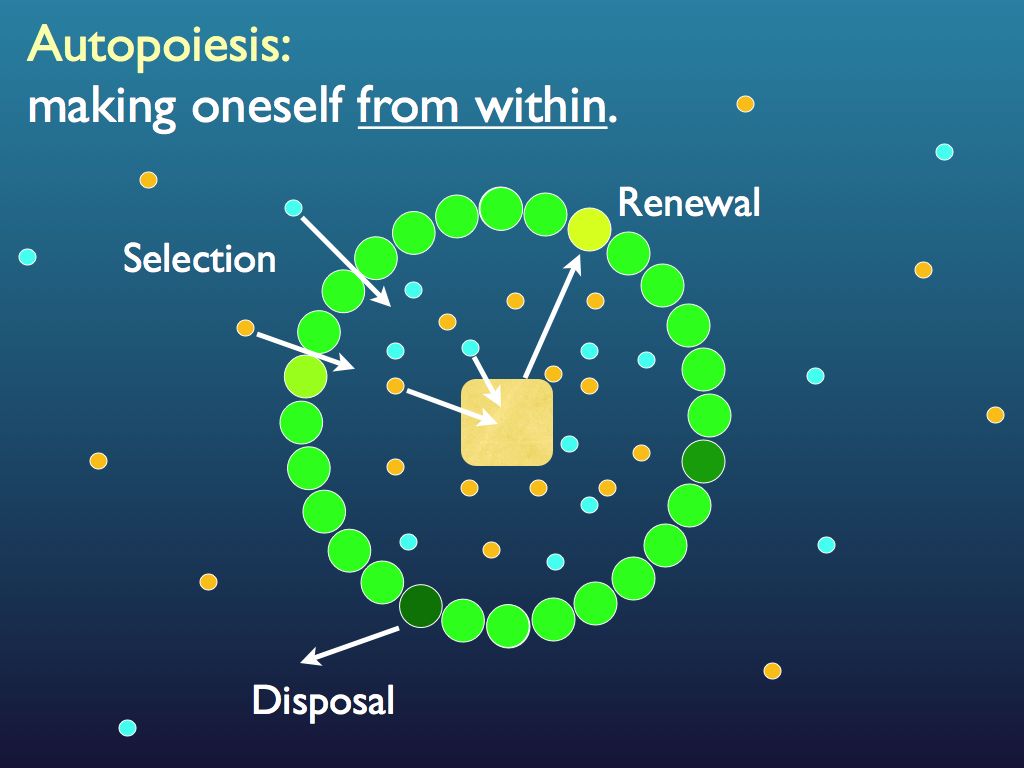
Whilst not autopoietic (due to the lack of a self-made boundary), the community does embody what seems to match the definition of a transcendent complex. It is multiply realisable (species can change and many are "redundant": as long as their significant functions are represented, the community is fine), in other words its components are treated as members of a functional equivalence class. It at least appears to exercise downward causation by selecting species for their functional contribution (via the rules for entry to the community). It has system-level (same level) functions which contribute to its own persistence.
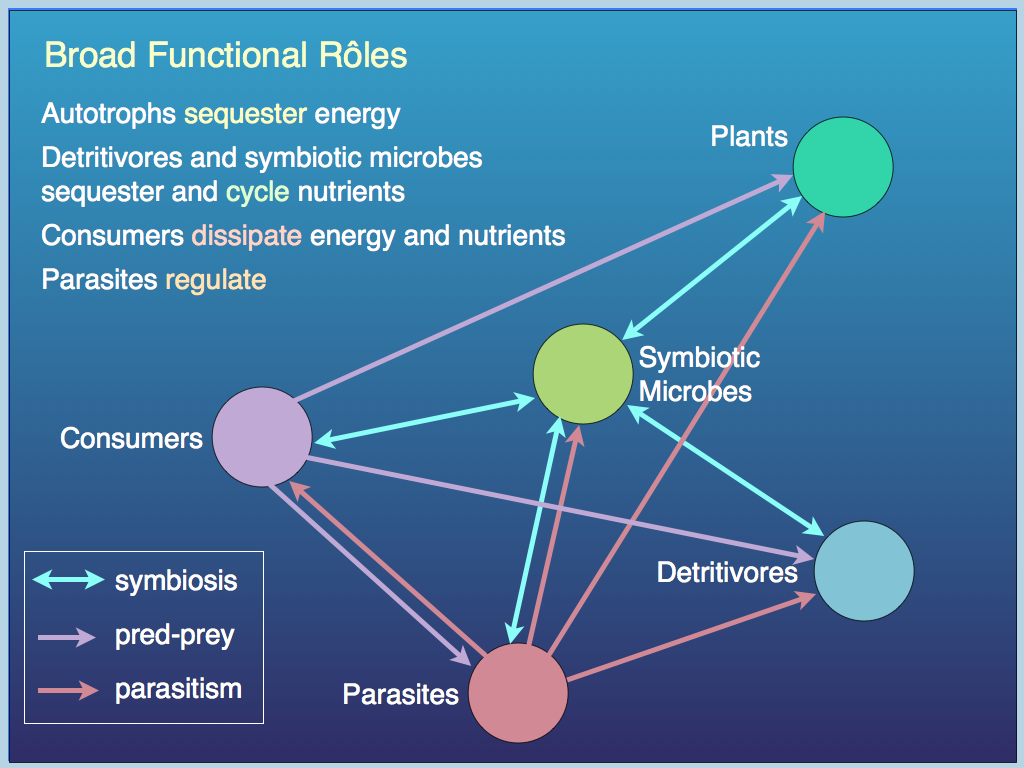
Different functional roles.
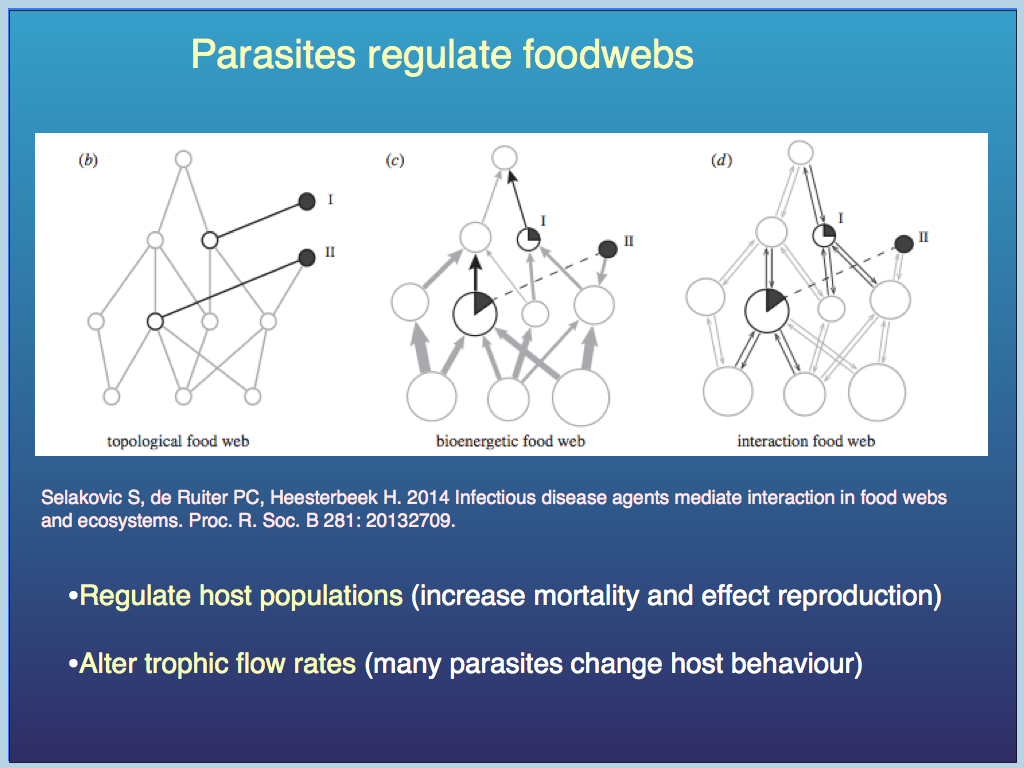
Food webs so dominate community ecology research that it is useful to briefly remind ourselves of the importance of the relatively neglected functional groups that are needed to make a viable ecological community. Let us start with the parasites. These are usually considered wholly negative , but (like many nasty things) they actually perform necessary and therefore valuable functional services.
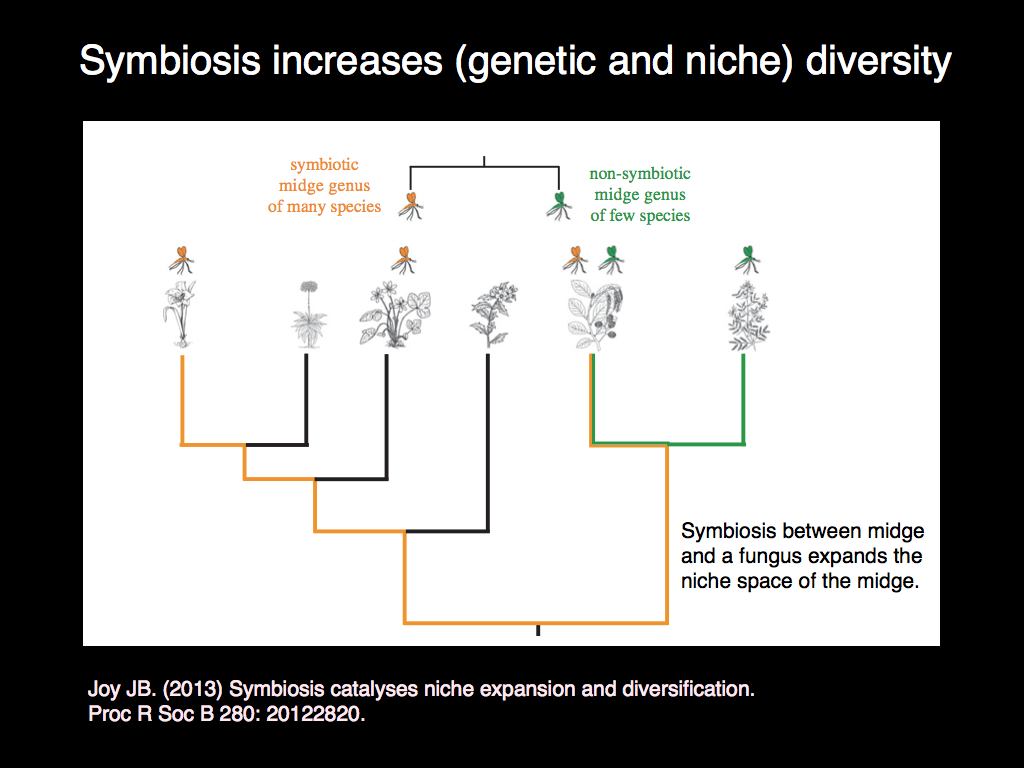
More embraced for their seemingly ultruistic contributions are the symbiotic species, though their apparent selflessness is very far from real (don't be misled by anthropocentric thinking). Symbiotes act as positive feedback connections in communities which constitutes one of the set of fundamental cybernetic control linkages. Having it increases the repetoire of community responses to change and in conjunction with the negative feedbacks provided by trophic and parasitic interactions, enables the system as a whole to, at least in principle, express all the attributes of an information processing system, perhaps even a Turing machine (though that is far from certain).
If you still need convincing that the community is an information processing (computation) system built from functional components in which the functions enable the set of general cybernetic control links and the community structure 'wires' them together to make a functional whole, then consider the emerging view of microbial communities.
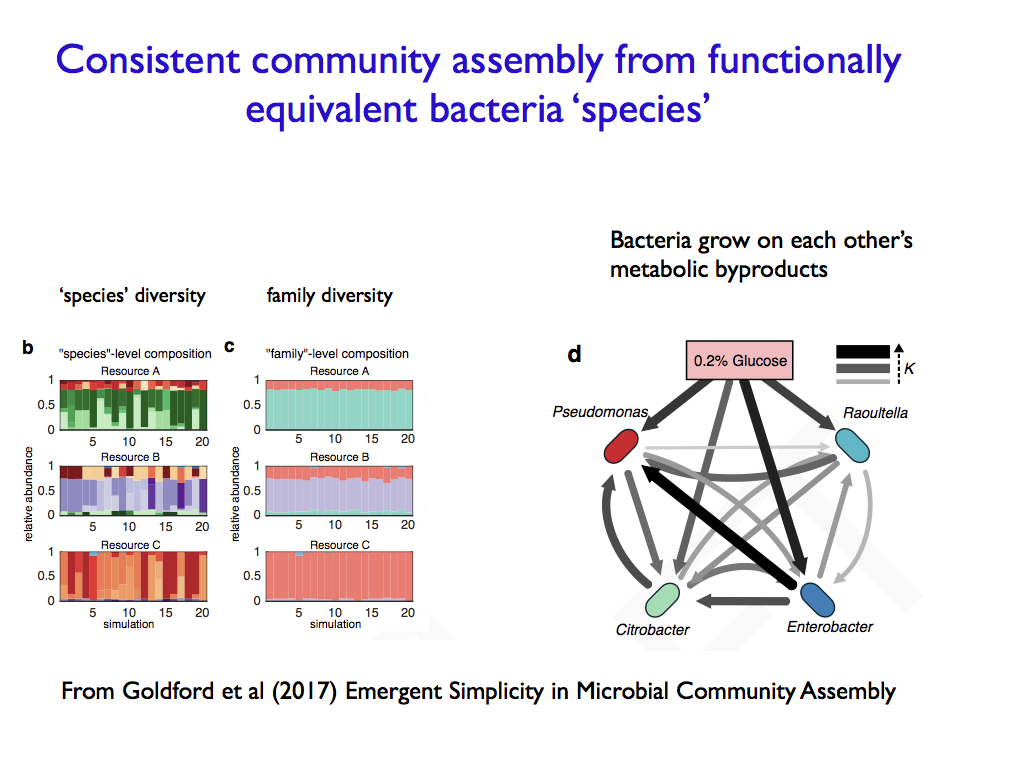
Some ecologists have moved away from the old-fashioned species based studies. Though species are not irrelevant, since they are still the foundation of population isolation that determines population dynamics, they are not fundamental to community ecology any more. Trait based ecology is gradually gaining ground, though the idea of a trait is still not usually well defined. Indeed, it is probably the difficulty of identifying functional traits that has hampered this progress. Among microbiologists, it is the idea of species that poses serious problems and traits, especially in terms of biochemical capacities, now take a prominent place in community models. Still, the interactions and their organisational and perhaps computational contribution is still largely neglected.
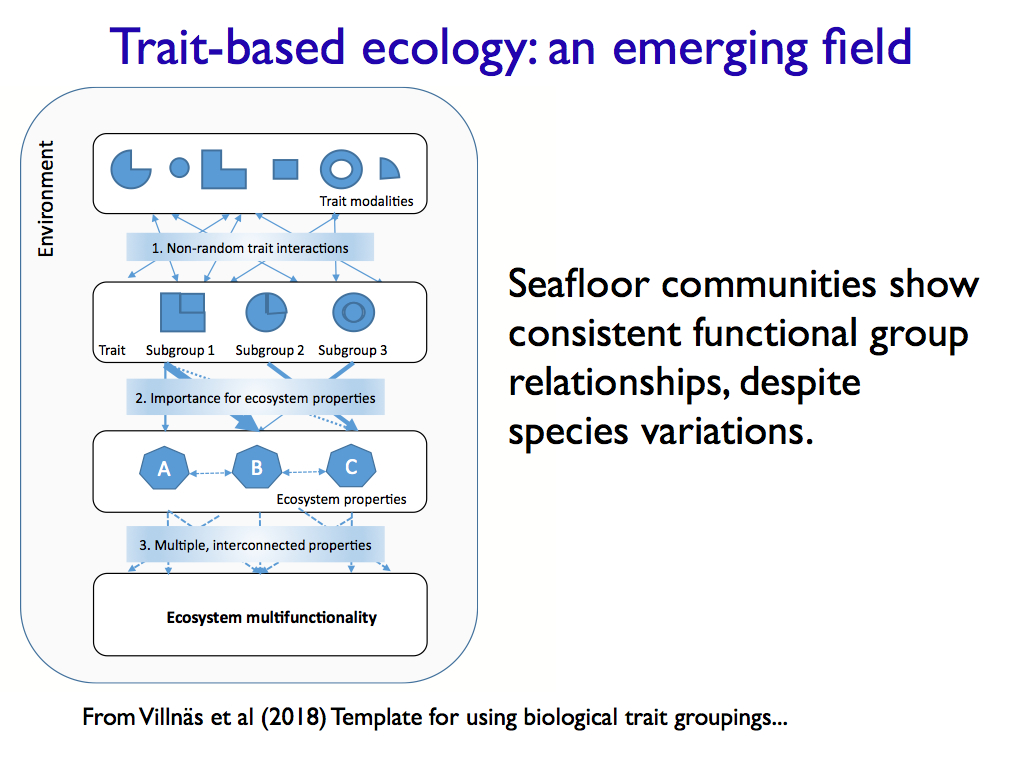
To summarise, I want to promote the idea of these interactions being crucial to community structure and find out how much the intangible information that they embody is responsible for the emergence of community level functions. I want to know if there really is a thing that contains and directs the community and computes its structural integrity (stability and robustness). In short, I want to know if the Ma of ecology is real. I illustrate these thoughts with the slides below.


These ideas were brought together in a paper defining ecological function (Farnsworth et al. 2017). It was inspired by both the frustration at the lack of rigour and realism in the very loose usage of the word function in ecology and also the great advantages of seeing ecological systems in terms of functions and relationships among them, rather than species (ecology has been hampered by its historical grounding in natural history).
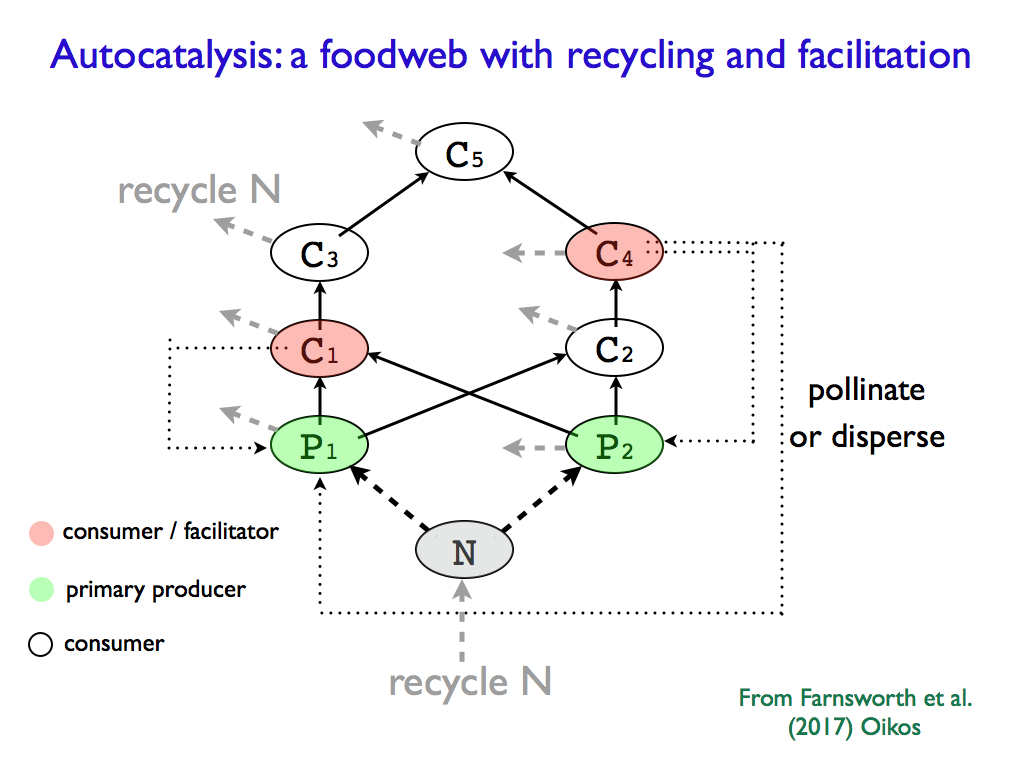
The diagram below, taken from the paper, shows a small, but functional ecological system in which two primary producer species (plants P1 and P2) compete and are consumed by two primary consumers (C1 and C2), themselves consumed by predators (C3, C4) which are prey to the top predator C5. Consumers C1 and C4 also function as seed dispersers (e.g. pollination or squirrels hiding nuts), i.e. facilitators. All the organisms produce nitrogen rich waste (a surrogate for all the nutrients needed by the plants). The identity of the species does not matter and many systems like this (though richer and more complicated in practice) can be found in nature, because it is the functions that define the system. Indeed species are substitutable and their role is to perform the required functions. The system is autocatalytic in the sense that each part of it promotes the functioning of each other part via its functional action as part of the whole. Obviously plants feed consumers and lower trophic consumers feed their predators, but some consumers promote the growth of the plant populations by facilitating their reproduction and providing accessible nutrients in waste for recycling. The higher predators function as regulators - important in keeping the system within the limits of its elasticity, preventing catastrophic instability. What is missing here are the recycling community of organisms and the regulating parasites, but this little system is enough to illustrate the point.
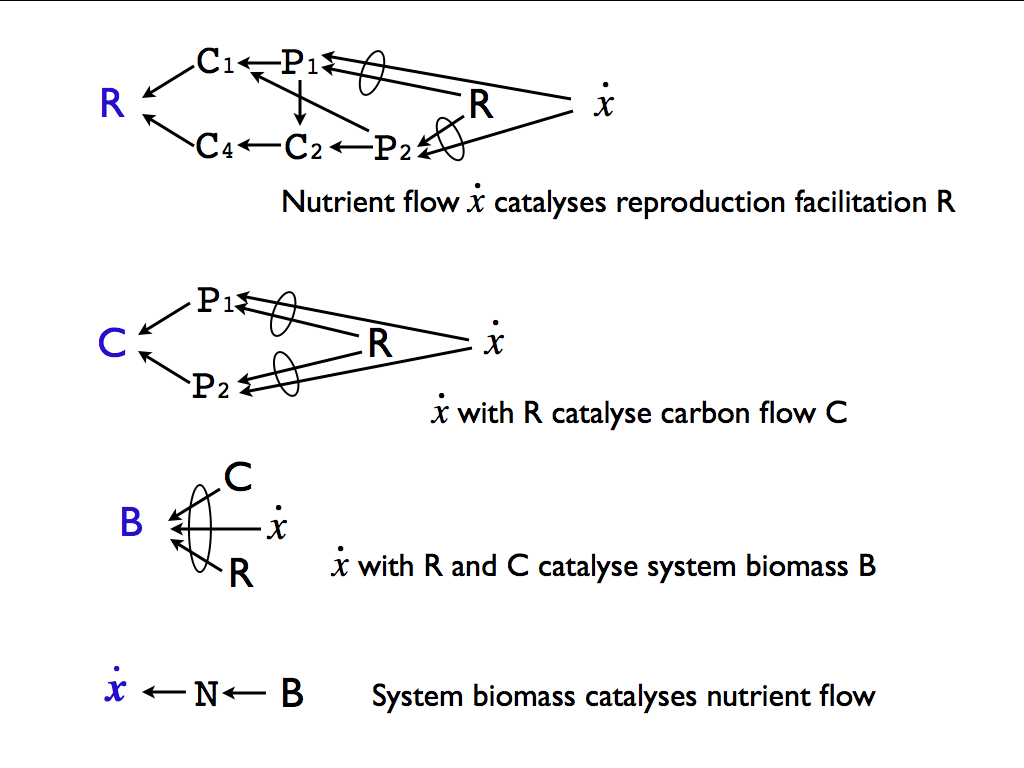
The ecological autocatalysis is emphasised by the dependencies illustrated in the next diagram (also from Farnsworth et al. 2017). We see first the flow of nutrients (x) [the dot on top denotes the time-derivative, so flow rate] as it promotes the function of facilitating reproduction (R), hence catalysing the plant production process. Then combining nutrient flow with this reproductive facilitation promotes carbon flow through the system (C). The carbon flow combined with R and nutrient flow promote the growth of biomass in the whole system (B), i.e. these flows catalyse growth throughout the system. Finally, growth of biomass increases the production nutrients (as waste) which increased the nutrient flow that enters as the starting point of the first diagram. Every part of the system makes a (catalytic) contribution to every other part and, more importantly, to the whole, with the result that biomass of the whole is increased. Indeed, it could be concluded that the function of the whole to which each component contributes is the production of biomass - and this was suggested as the overall master function of an ecological community, and in fact, of life overall.
Farnsworth, K.D.; Albantakis, L.; Caruso, T. (2017). Unifying concepts of biological function from molecules to ecosystems. Oikos, doi: 10.1111/oik.04171.
It is said that the essence of a pot is the space it contains; the essence of a room is the space it encloses: space completes the form and gives it meaning.
Japanese culture names this ‘Ma’: the empty space that gives things meaning. It is exemplified with the typical economy of Japanese art in the beautiful screen printing shown above.
My question is this: can we apply the principle of Ma to better understand what an ecosystem is? Of course, the answer is going to be yes. Let us see why.
Structure
First, rooms and pots, etc. are physical objects that are what they are because of the specific placement of material in space and time: they are geometric structures of material substance.
When it comes to living nature, ecologists typically survey a natural community, count the species and the numbers of organisms of each and then call this ''community structure'' (see the biodiversity indices page).
A catalogue of species present and their relative abundances is not what most people would call structure.
This is more like it, though it is a human artifact, the Forth Bridge is an ordered set of material components in which the order (placement of each in relation to the others) creates a whole. It is an example of the general class of engineering structures.
All members of this class share the following:
a) an underlying topology determines which components are connected to which and by what connecting points (be they connectors, nuts and bolts or points for welding etc.);
b) a geometry in which the topology is expressed in physical space;
c) a physical form, which instantiates the design using physical components arranged and interconnected in the real geometry. In the example illustrated above, a simple radio receiver is first specified as a topology connecting specific functional components in a particular way, this is then extended in space using printed circuit board tracks to make material the (electrical) connections and then the assembly, including the physical components, completes the structure. Please note that at every one of the three stages, organising (constraining) information is added to specify a particular configuration of parts, not any configuration, just this one will work as a radio. Is it possible that the same can be said for an ecological community?
We can at least conceive of a model of such using the same principles, as this example (above) from Ernstson et al (2010) shows.
In general terms, the idea is that the information specifying the links of interaction among the component organisms constitutes the information component of the structure of the community. The other component is its physical material, just as in the Forth Bridge.
Embodied Information and Constraint
The set of interactions is information because it is specific to the community: recall that among the component we can have a large number of possible configurations of links and specifying just one from among all these is equivalent to a large reduction in the system's information entropy, which is equivalent to it gaining a large amount of information (see diagrams below and see the entropy page for a fuller explanation). Note that usually the precise configuration of links and components is not important to the community as a whole and in fact changes through time in a way that may as well be random. This is redundancy that is allowed because it is the functional relationships that matter to the community, not every detail of how they are created. Indeed, as we shall see shortly, the specific identity of organisms and even of species is not necessarily important to the community because it is only the functions they perform that matters at the community level. As such, the information is less than that of any exact specification of all interactions at a fixed point in time. The components, with their functions (based on functional traits) form a functional equivalence class for the community. This information is embodied at the system level as the set of interactions and it is functional because the system as a whole gains capacities from it, not least of which is the ability to self regulate and self-repair (summarised phenomenologically as 'stability' and 'robustness').
Taking the specific and very simple example (below), we can look at the difference between a network of interacting species that is a) simply a uniform interconnection (as for example in a community of generalist (opportunist) predators and prey, where the only 'rule' is that if a predator of any species meets a prey of any species, then the outcome is not good for the prey) and b) a community of specialists, in which each predator fastidiously only chooses to eat a specific prey species, leaving all the others. The generalist community has a network that is uninformative. We calculate its entropy using Shannon's formular, in which the p values are the probrabilities that a connection (predation incident) is made. In the uninformative community all the ps equal 1 and we get a system entropy of 4.75. But in the community of specialists, most of the ps are zero and the system entropy is the much smaller 1.585. That means that this specialist community has embodied information at the system level, an amount equal to the entropy of the most uninformative community possible, minus its own entropy. That works out as 3.17 bits of information in this little example.
Now here is a real life example: the symbiosis network of interactions between flowering plants and their pollinators. I did not work out the entropy of this system, but given the data, we could do that. The point emphasised below is that the network is very far from uniform, uninformative. There are strong associations and very weak or near p=0 ones and this results in a distinct patter of interactions, which even contains a higher order structure, as shown in the graph of pollinator species against plant species (coloured squares showing strong interactions). This higher level structuring is revealed as clusters coinciding with the colours.
By far the majority of ecological networks studied professionally are trophic networks - foodwebs. My former colleauge Axel Rossberg has studied the effect and importance of non-random preferences of predators for particular species, building realistic models based on the idea of trophic trait matching. The idea is that a predator (a) is more likely to take prey (b) when the predator has traits that complement those of a prey. Taking a very obvious example, they both live in the same sort of habitat.
So, we may reasonably think there is a general pattern here.
The information caused by all the particular selections making up the non-random network of interactions is embodied at the community (system) level and is responsible for functions that arise from that level. We should recall what is really meant by function here. It is a capacity of an organisational level to contribute to the capacity of the next higher level. In Farnsworth et al. 2017,
we explain what this means at the level of ecological community.
Whilst not autopoietic (due to the lack of a self-made boundary), the community does embody what seems to match the definition of a transcendent complex. It is multiply realisable (species can change and many are "redundant": as long as their significant functions are represented, the community is fine), in other words its components are treated as members of a functional equivalence class. It at least appears to exercise downward causation by selecting species for their functional contribution (via the rules for entry to the community). It has system-level (same level) functions which contribute to its own persistence.
Different functional roles.
Food webs so dominate community ecology research that it is useful to briefly remind ourselves of the importance of the relatively neglected functional groups that are needed to make a viable ecological community. Let us start with the parasites. These are usually considered wholly negative , but (like many nasty things) they actually perform necessary and therefore valuable functional services.
More embraced for their seemingly ultruistic contributions are the symbiotic species, though their apparent selflessness is very far from real (don't be misled by anthropocentric thinking). Symbiotes act as positive feedback connections in communities which constitutes one of the set of fundamental cybernetic control linkages. Having it increases the repetoire of community responses to change and in conjunction with the negative feedbacks provided by trophic and parasitic interactions, enables the system as a whole to, at least in principle, express all the attributes of an information processing system, perhaps even a Turing machine (though that is far from certain).
If you still need convincing that the community is an information processing (computation) system built from functional components in which the functions enable the set of general cybernetic control links and the community structure 'wires' them together to make a functional whole, then consider the emerging view of microbial communities.
Some ecologists have moved away from the old-fashioned species based studies. Though species are not irrelevant, since they are still the foundation of population isolation that determines population dynamics, they are not fundamental to community ecology any more. Trait based ecology is gradually gaining ground, though the idea of a trait is still not usually well defined. Indeed, it is probably the difficulty of identifying functional traits that has hampered this progress. Among microbiologists, it is the idea of species that poses serious problems and traits, especially in terms of biochemical capacities, now take a prominent place in community models. Still, the interactions and their organisational and perhaps computational contribution is still largely neglected.
To summarise, I want to promote the idea of these interactions being crucial to community structure and find out how much the intangible information that they embody is responsible for the emergence of community level functions. I want to know if there really is a thing that contains and directs the community and computes its structural integrity (stability and robustness). In short, I want to know if the Ma of ecology is real. I illustrate these thoughts with the slides below.
These ideas were brought together in a paper defining ecological function (Farnsworth et al. 2017). It was inspired by both the frustration at the lack of rigour and realism in the very loose usage of the word function in ecology and also the great advantages of seeing ecological systems in terms of functions and relationships among them, rather than species (ecology has been hampered by its historical grounding in natural history).
The diagram below, taken from the paper, shows a small, but functional ecological system in which two primary producer species (plants P1 and P2) compete and are consumed by two primary consumers (C1 and C2), themselves consumed by predators (C3, C4) which are prey to the top predator C5. Consumers C1 and C4 also function as seed dispersers (e.g. pollination or squirrels hiding nuts), i.e. facilitators. All the organisms produce nitrogen rich waste (a surrogate for all the nutrients needed by the plants). The identity of the species does not matter and many systems like this (though richer and more complicated in practice) can be found in nature, because it is the functions that define the system. Indeed species are substitutable and their role is to perform the required functions. The system is autocatalytic in the sense that each part of it promotes the functioning of each other part via its functional action as part of the whole. Obviously plants feed consumers and lower trophic consumers feed their predators, but some consumers promote the growth of the plant populations by facilitating their reproduction and providing accessible nutrients in waste for recycling. The higher predators function as regulators - important in keeping the system within the limits of its elasticity, preventing catastrophic instability. What is missing here are the recycling community of organisms and the regulating parasites, but this little system is enough to illustrate the point.
The ecological autocatalysis is emphasised by the dependencies illustrated in the next diagram (also from Farnsworth et al. 2017). We see first the flow of nutrients (x) [the dot on top denotes the time-derivative, so flow rate] as it promotes the function of facilitating reproduction (R), hence catalysing the plant production process. Then combining nutrient flow with this reproductive facilitation promotes carbon flow through the system (C). The carbon flow combined with R and nutrient flow promote the growth of biomass in the whole system (B), i.e. these flows catalyse growth throughout the system. Finally, growth of biomass increases the production nutrients (as waste) which increased the nutrient flow that enters as the starting point of the first diagram. Every part of the system makes a (catalytic) contribution to every other part and, more importantly, to the whole, with the result that biomass of the whole is increased. Indeed, it could be concluded that the function of the whole to which each component contributes is the production of biomass - and this was suggested as the overall master function of an ecological community, and in fact, of life overall.
Farnsworth, K.D.; Albantakis, L.; Caruso, T. (2017). Unifying concepts of biological function from molecules to ecosystems. Oikos, doi: 10.1111/oik.04171.